 I attended the annual meeting of the Society for Vertebrate Paleontology (SVP) last week. This is the yearly meeting where those studying anything with a backbone, from fish to furballs get together to talk about what they have been doing and see what others have found (although admittedly, most of the press goes to dinosaurs). Over the next few posts, I will share brief snippets of what I learned. If you read anything that sounds particularly interesting to you and would like me to discuss it in more detail, just let me know in the comments section and I can expound on the topic.
I attended the annual meeting of the Society for Vertebrate Paleontology (SVP) last week. This is the yearly meeting where those studying anything with a backbone, from fish to furballs get together to talk about what they have been doing and see what others have found (although admittedly, most of the press goes to dinosaurs). Over the next few posts, I will share brief snippets of what I learned. If you read anything that sounds particularly interesting to you and would like me to discuss it in more detail, just let me know in the comments section and I can expound on the topic.
So what does a scientific meeting like this look like? There is always a dealers’ room, where you can find an array of book publishers, such as Indiana Press and CRC Press, and supply companies, such as Paleo-Tools and Bone Clones, and many others all hawking their wares. You will meet artists such as Luis Rey, learn about what’s going on at different publications, such as PLOS One, and upcoming meetings. There are the business meetings, social events, and award banquets. There are field trips to local paleontolgical sites (this meeting was in Los Angeles, so the La Brea tar pits and the Los Angeles Natural History Museum were highlights). There is also the chance to converse with paleontologists from all over the world, a venue wherein colleagues can talk face-to-face and forge new connections, where students can get their feet wet presenting at an international conference and talk to students of other professors to learn about potential graduate and post-graduate opportunities, which professors make good mentors and which ones to avoid. And of course, there are the talks and poster sessions, where you can hear about the research people are doing right now. If you want to know the current state of the field, this is the place to go. Every day for four days, three sessions run concurrently, each covering 16 talks a day, plus over 120 posters are presented every day. By the end of which, if you haven’t found enough new information to send your brain into overload, you simply haven’t been trying. This is also a relatively small meeting, with only 1500 or so attendees, unlike some scientific societies which have meeting attendances over 20,000.

Bosch Institute Neuroscience meeting, 2007. This is a bad design for meetings. Please, just stop.
Before I get into a short recap of the talks I attended (there were many more I would have liked to attend, but I have not yet perfected cloning myself), I will digress a moment for a quick comment about the meeting room setup. My first thought upon seeing the room arrangement was Happy day! All the meeting rooms are right next to each other; not on a different floor, on the other side of a giant building, in another building, or in another facility entirely, unlike some other meetings I’ve attended. This makes popping from one session to another to see different talks much easier. The downside to this is that hundreds of other people all have the same idea and are trying to go through the same hallway, trying to get around the other attendees who are chatting with friends and colleagues. For those of you that are accustomed to New York subways, this is no big deal, but for some of us, it can be a bit claustrophic as personal space shrinks to microscopic proportions. The other rant I have is that, after all this time, they still have not learned how to lay out a room for the talks. They insist on preparing the rooms as though everyone will calmly file in before the talks and sit there until the break, providing long rooms with the speaker at one end and two long columns of interlocking chairs set in rows up to 20 seats long. however, this is a scientific meeting with many short talks. people stream in and out constantly. So what happens is that all the edge seats are quickly taken and great, yawning chasms of emptiness are left in the center. Why no one ever thinks to place the speakers in the middle of a long wall, with many columns of seats with no more than 6-7 seats to a row, allowing ample space for people to move, is beyond me. They might not be able to put as many seats in, but that hardly matters if no one can get to the seats in the first place. Now on to the talks!

Sauropod nest, by Mike Ellison
In the first session, all the talks dealt with the problems of ontogeny in interpreting the fossil record. Ontogeny, how an animal grows from fertilized egg to old age, can cause several problems in the fossil record. When all you can see is a fossil, it can be very hard to tell if you are looking at an adult or juvenile. Many animals can change so much during development that the juveniles can look like completely different species. Hans Larsson presented about a possible way to figure out rough ages of an animal by looking at how different bones in the skull covaried, or how they changed shape in relation to each other as they grew. Jack Horner warned against assuming an animal is full-grown unless you have solid evidence of that from the bones across the skeleton and Mark Goodwin showed how difficult that can be with pachycephalosaurs, but Holly Woodward found that at least some dinosaurs reached sexual maturity long before they reached “adult” size and in fact, never stopped growing, so it is important to keep in mind just how one defines “adult.” David Evans showed the problems of using juveniles and adults in the same phylogenetic analysis, in that the relationships of species identified using juveniles were very ambiguous and unstable, often showing up as more ancestral than an adult of the same species, sometimes substantially so. John Scannella found that using just one juvenile specimen of Triceratops in a phylogenetic analysis resulted in that specimen not being identified as a ceratopsian at all, but if a sufficient number of juvenile specimens of different ages were included, they all fell out together in their expected relationship. This indicated the ontogeny problem can be dealt with if you have enough samples, as well as indicating what characters are newly evolved versus being evolutionarily stable. Zachary Morris extended this to show that not all individuals grow up the same way, there was no set pattern of development that all individuals within the same species followed to adulthood, so trying to order specimens into an ontogenetic sequence requires a large sample size. Also, contrary to an earlier research that got a lot of press, Torosaurus is probably not the same species as Triceratops. Robert Reisz showed very fast growth in prosauropod embryos and P. Martin Sander estimated that sauropods could grow to sexual maturity in 16 years, attaining 90% of their maximum size within 32 years. This comes out to an average growth rate of 4 kg/day, which is similar to modern large mammalian herbivores, but he cautioned this was likely overestimating the ages and underestimating the growth rates. Ken Dial discussed the role of predation in the development of flight, with evidence indicating that if predation pressure was relaxed and there was sufficient food resources, birds commonly became flightless.He made the interesting point that everything is a transitional environment; that to truly understand an organism,one has to examine all aspects of their habitat and you can’t afford to get to narrowly focused. Denver Fowler found that animals occupying different niches at different ages were more susceptible to extinction. Caroline Stromberg found that, contrary to popular belief, the development of grasslands didn’t actually correlate very well with high-crowned, hypsodonty teeth.
Wikipedia.org
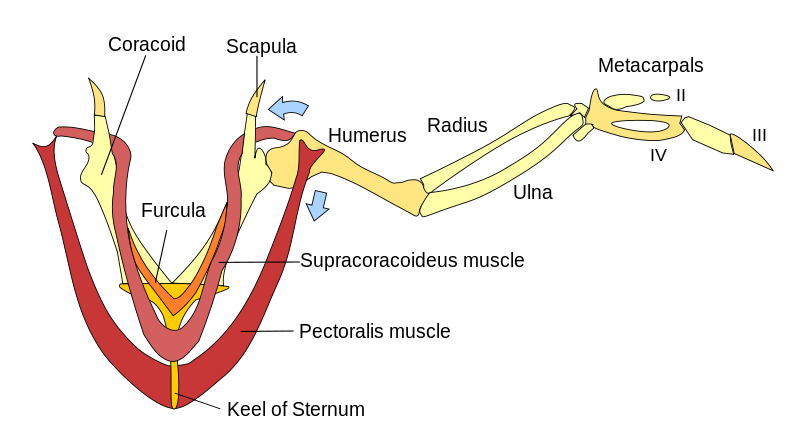
Sandy Kawano looked at how locomotion changed between fin and feet and found that the total forces between front and hind limbs was fairly constant, but acceleration came from the pectoral fins at first, but as the hind limbs developed, more of that motive force came from the hind legs, with the front legs taking on more of a regulatory role guiding maneuverability. Karen Sears found that limb evolution became more modular, in that different parts of the limb evolved at different rates and times, so that each part could be considered as evolving relatively independently, with higher levels of evolution near the ends of the limbs. Paul Sereno discussed the evolution of the coracoid bone in the shoulder and how its loss in almost all mammals created a more flexible and faster, but weaker joint. He opined this is why bats don’t grow to the size of birds or pterosaurs and that it allowed dinosaurs to get bigger and carry more weight than mammals.
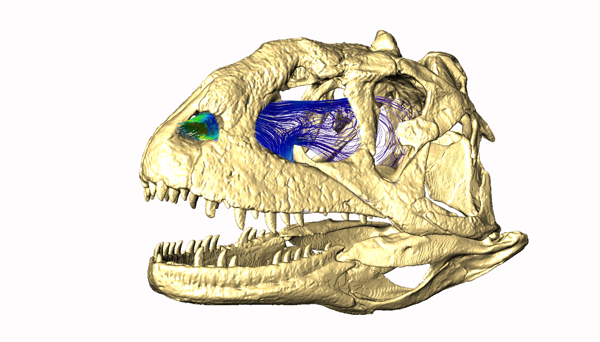
Model of air flow through the nasal passages of Majungasaurus. Image and work by Jason Bourke
Ali Nabavivadeh reported on his studies on the evolution of jaws in ornithischian dinosaurs, finding they talked the problem in a variety of different ways. Lucas Spencer noticed that all early ornithischians were in Gondwana and that dispersal explains more of their biogeography than vicariance (speciation via the emergence of physical barriers). Mark Loewen reported on the biogeography and phylogeny of ankylosaurs, while Victoria Arbour discussed the evolution of the ankylosaur tail club. Philip Currie reported on the smallest known articulated ceratopsid fossil, while Andrew Farke reported on ceratopsian biogeography. Thomas Carr found evidence to support anagenesis (direct ancestor-descendant relationships) in some tyrannosaurs, but not all, while Brandon Hedrick found that fossilization processes made one species of psittacosaurs look like three different ones. Jordan Mallon found little evidence in pachycephalosaur skulls to indicate they lived in upland areas, as is often stated, and Jason Bourke did some very nice computer modeling showing how turbinates (structures within the nasal passages often associated with endothermy, aka warm-bloodedness) work to channel air through nasal passages,using that to find support for respiratory turbinates in pachycephalosaurs.
This all only recounts brief snippets of the talks I attended on the first day and doesn’t include the many posters I looked at. I will cover Day 2 in my next post. If you want more information on any of these snippets, leave a comment and I will be happy to expound upon them.


[…] Day 1 at SVP (paleoaerie.org) […]
[…] in Arkansas and the dinosaur “Arkansaurus,” and went to the annual meeting of the Society of Vertebrate Paleontology. We looked at geologic time and started a series on dinosaur misconceptions. We also had several […]