Mystery Monday

This week’s fossil is a pretty little coiled animal. Can you guess what it is?
Taking a Bite Out of Fossil Friday
My apologies for getting this out so late. But now that we’re here, could you tell what the mystery fossil for this week was? It was a very unusual creature. 

Peafowl, male and female. Wikimedia.
The creature in the picture is a representative of an early shark called Falcatus. It was a very strange shark, in which the males had a forward-pointing spike on its head. This is one of the earliest examples of sexual dimorphism in the fossil record. Sexual dimorphism is when the male and females look different. Humans are sexually dimorphic in a variety of ways, but the classic example is the peacock with its extravagant tail. They are so dimorphic that many people do not even realize the term peacock only applies to the male of the peafowl species. The females are called peahens.

Falcatus reconstructed. Wikimedia.

Ozarcus. The fossil itself. AMNH.
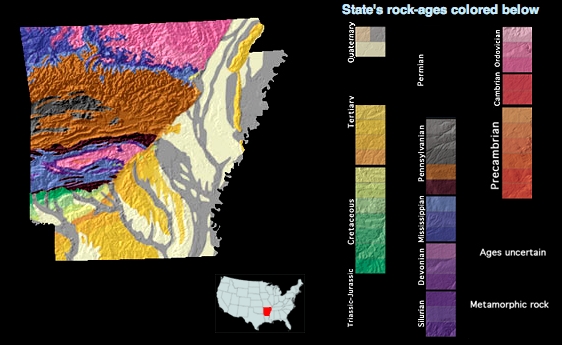
Falcatus was chosen this week because of an interesting shark fossil found in 325 million year old limestone near Leslie, AR. The fossil was recently published and got a lot of press. Pictures of the fossil itself show what appears to be little more than a couple of lumpy concretions stuck together. There is little there that resembles anything like a shark. At least, until you look inside. A concretion is an inorganic structure consisting of layers of minerals that have been precipitated around a central core, much like a rocky onion. Oftentimes, a fossil lies at that core and served as the basis upon which the mineral precipitation got started. So one never knows what one will find cracking open a concretion. It may be nothing more than concentric mineral bands, or it may be an exquisite fossil preserved for the ages. In this case, the scientists got lucky. Not only was there an exquisite fossil, but one in which few had ever seen before: the skeletal structure of an early shark’s gill basket.
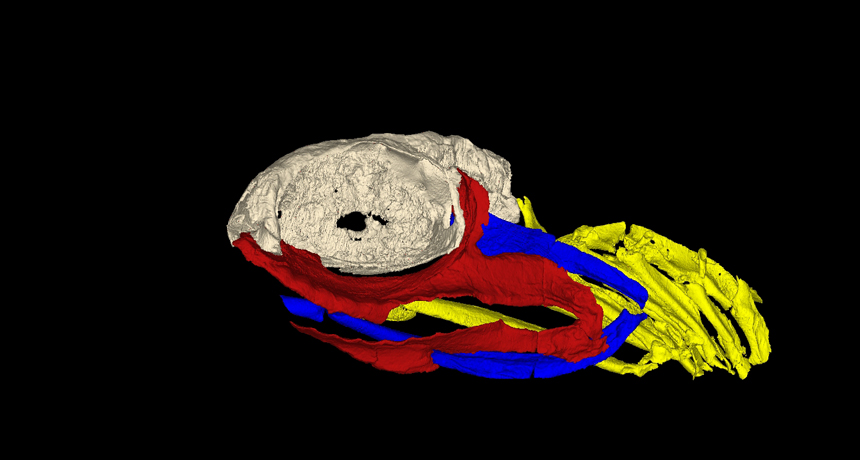
A 3D reconstruction of the skull of Ozarcus mapesae. The braincase is shown in light grey, the jaw is shown in red, the hyoid arch is shown in blue, and the gill arches are shown in yellow. ©AMNH/A. Pradel
The researchers named the fossil Ozarcus mapesae, for the Ozarks in which it was found and for Royal and Gene Mapes, geology professors at Ohio University. The Mapes are also paleontologists and have collected a large number of fossils, one of which happened to be this curious-looking concretion. Small teeth on the exterior pointed to possibly more interesting material inside, so they CT-scanned it, which revealed the remains of the head of the shark. But to get better detail, they had to use a synchotron. This technology is new and expensive enough that it was not possible to look at the fossil this way until recently (a good example of why we preserve fossils in museums, you never know when future technology will allow examination in ways never thought of before).
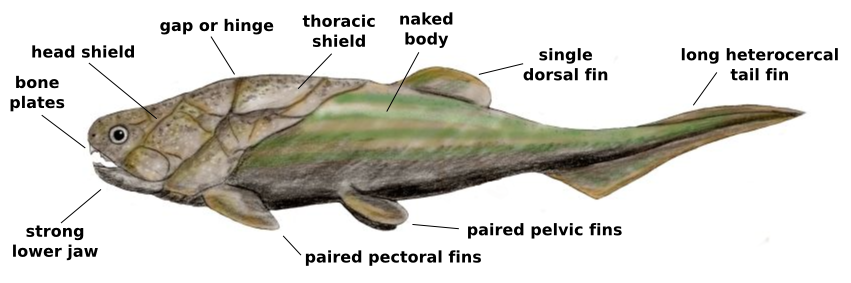
Placoderm anatomy. Wikimedia.
The gill basket goes by many names, the branchial basket, branchial arches, gill arches, etc. but they all refer to the skeletal supports for the gills. To the shark, they are important for holding up the gills and forming the path for water to flow over the gills so the shark can breathe. For us, they are important because the arches evolved into a variety of structures, primarily the jaws and hyoid, a small bone in the throat to which the back of the tongue is attached. Because of the importance of jaws, the evolution of those structures has been a big topic of interest. The traditional view has been that the first jawed fish, the placoderms, evolved into the early precursors of the sharks, which then evolved into the early bony fish, the osteichthyans. The sharks then were expected to have a more primitive structure than bony fish. This story makes sense, given that the order of appearance in the fossil record pretty much matches what we would expect and the skeletal structures look more primitive in sharks than bony fish. Of course, despite the common view that sharks are relics of a bygone age, they have had over 400 million years to evolve after they split off from bony fish. How likely is it that they would have retained such ancestral characteristics for all this time? To answer that question, we can look at the fossil record to tell us what they were like at the earliest stages.
 The big problem with examining the fossil record of sharks is that sharks are chondricthyans. They do not make bone. Other than the teeth, the skeleton is supported by a simpler set of calcium phosphate crystals. Unlike bone, which has a very structured arrangement of crystals and connective tissue, the bones in sharks are made of cartilage and a haphazard set of disorganized crystals, which fall apart shortly after the animal dies. Thus, finding any fossils of sharks that contain more than the teeth is extremely rare. Finding ones in which everything is still in place is almost impossible. Fortunately, over a long enough period of time, even the almost impossible will happen eventually. That is the thing with large numbers and vast amounts of time, our perception of what is unlikely doesn’t really work. For instance, given a 1 in a million chance that a tweet on Twitter will have something requiring the security team to deal with, that still gives them 500 tweets every single day. That “almost impossible” fossil was found with Ozarcus. This fossil provided our first look at what the throat of a primitive shark actually looked like.
The big problem with examining the fossil record of sharks is that sharks are chondricthyans. They do not make bone. Other than the teeth, the skeleton is supported by a simpler set of calcium phosphate crystals. Unlike bone, which has a very structured arrangement of crystals and connective tissue, the bones in sharks are made of cartilage and a haphazard set of disorganized crystals, which fall apart shortly after the animal dies. Thus, finding any fossils of sharks that contain more than the teeth is extremely rare. Finding ones in which everything is still in place is almost impossible. Fortunately, over a long enough period of time, even the almost impossible will happen eventually. That is the thing with large numbers and vast amounts of time, our perception of what is unlikely doesn’t really work. For instance, given a 1 in a million chance that a tweet on Twitter will have something requiring the security team to deal with, that still gives them 500 tweets every single day. That “almost impossible” fossil was found with Ozarcus. This fossil provided our first look at what the throat of a primitive shark actually looked like.
Let’s have a bit of background to cover what we have known of early jaw evolution up to this point. Placoderms, armored fish from the early Paleozoic Era, were the first animals with jaws. The jaws themselves appear to develop from the first gill arch, according to a lot of embryological studies on modern animals. 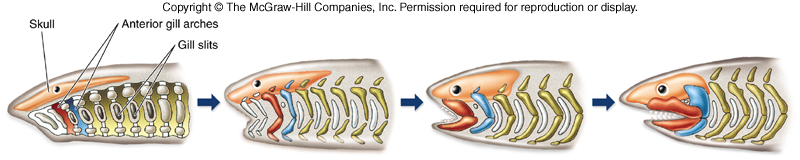
The studies haven’t really answered where the bone came from though. In placoderms, it is pretty clear the armor came from modifications of the dermis, the basal layer of the skin. But they also have internal bone forming their skeleton. Modern sharks have no bone other than teeth and bony fish have jaws made from that dermal bone. In addition to the origins of the bone, there is the matter of how the bones are attached to the skull. The upper jaw is formed by a embryological structure called the palatoquadrate (the top part of the first gill arch), so named because bones called the palatine and quadrate form from it. The bottom jaw forms from what is called Meckel’s cartilage (the bottom part of the first gill arch). In modern fish, the palatoquadrate is braced against the skull only at the front, with the back unattached. The jaw joint itself is attached to a bone called the hyomandibular, which forms from the second gill arch. The hyomandibular acts like a swinging pivot, allowing the jaw to open very wide. When the jaw joint gets pulled forward, the back of the upper jaw can move down, using the point where the front part is braced as the pivot point. Sharks take this to an extreme, not bracing the upper jaw on anything at all, with the jaws attached solely by the hyomandibular bone (the “hyostylic” joint) allowing both the upper and lower jaw to move forward when they open their mouths. So when you see that shark opening its jaws and it looks like they are coming right out at you, they really are. For a great example of a hyostylic jaw joint, check out the goblin shark.
Unfortunately, when we look at the earliest fossil sharks and bony fish, both of them show a jaw in which the upper jaw is braced against the skull in both front and back (the “amphistylic” joint). So it doesn’t tell us much about how the sharks fit into the sequence. One might say that it seems logical to think the bony fish came first, loosening the jaw in the back and then the chondrichthyans took this one step farther. But surely, others might say, the fact that bony osteichthyans have a more advanced bony skeletal structure means they would have to have come later, right? Here again, the fossil record doesn’t help here because the earliest representatives of both groups appear close enough in time that it cannot be strongly stated which came first.
That is where things were, until 2013, when the fossil record started answering these questions. A placoderm called Entelognathus was published in 2013. This fossil was of great interest because it showed the structure of the jaws in great detail. Entelognathus had a jaw that looked very much like an osteichthyan. This fossil was 419 million years old, so it was likely too late to be ancestral to either chondrichthyans or osteichthyans, but old enough to be very close to the ancestral form. What Entelognathus tells us is that the bones forming the jaws in placoderms was already like those seen in modern bony fish, indicating that sharks would have started out with bone, but lost it during their early evolution. Of course, since sharks don’t have bone, this was hard to demonstrate on sharks, so the question was still unsettled.
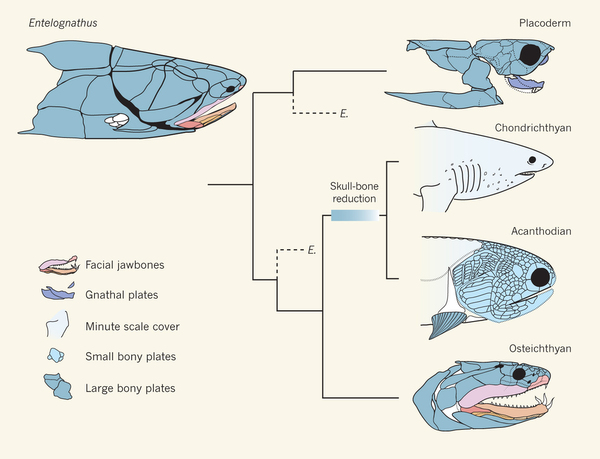
What Entelognathus says about fish relationships. Palaeontology: A jaw-dropping fossil fish. Matt Friedman & Martin D. Brazeau Nature 502, 175–177 (10 October 2013) doi:10.1038/nature12690
Outside of the actual jaws themselves, there are all the other support structures around the jaw, such as the hyoid bone. Many of these structures are formed from the next few gill arches (earlier jawless fish had at least seven gill arches, so using a few to make the jaws and throat still leaves plenty for gills). The skeletal structures in bony fish that support the gill arches form a fairly simple chevron, or wide v shape, whereas the sharks have a slightly more complicated structure. But if finding fossils of shark skeletal structures is rare, finding one with tiny gill arch supports still in position is almost impossible. There is where Ozarcus comes in, because it is here that the almost impossible becomes reality. Once the researchers were able to see inside the fossil with the synchotron, they were able to see preserved gill arch supports in position. That position resembled the simple chevron shape of modern osteichthyans.
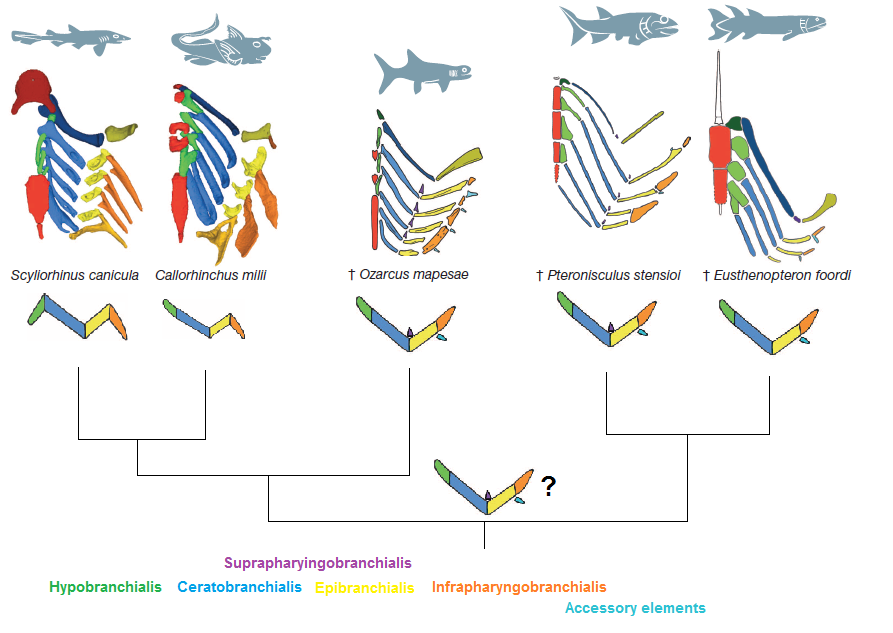
Gill arch supports in Ozarcus are similar to bony fish (right) and not the condrichythyans (left). Pradel A., Maisey J.G., Tafforeau P., Mapes R.H. and Mallatt J. (2014) A Palaeozoic shark with osteichthyan-like branchial arches. Nature DOI: 10.1038/nature13195
Thus, between Entelognathus and Ozarcus, we can confidently assess the development of jaws as having started in placoderms with primitive, osteichthyan jaws. We have evidence of the bony origins of the jaws from Entelognathus and evidence from the gill arches from Ozarcus, so we now have plenty of evidence to strongly support the claim that the chondrichthyans are not in the evolutionary pathway to modern jaws at all. They are an offshoot that actually lost bone to form a more flexible skeleton for some reason. Whether it was mechanical advantage for their lifestyle, such as increasing flexibility, or a reduction in mineral storage, who can say.
This new view of jaw evolution demonstrates that the common view of sharks as being evolutionary relics is wrong. Far from being primitive, they evolved throughout the millenia to the modern sharks we see today, changing the shape and development of their jaws and throat as they evolved from the primitive condition seen in early bony fish into the efficient predators of today. But then, they have been separated from the lineage that led through bony fish up through the early tetrapods all the way to us for over 400 million years. Why would anyone think that in all that time, they stayed still evolutionarily? Even if they look much more similar to their ancestors than we do, they have evolved too. Nothing stays still.
Website Review and the Misconception of a Theory
In a quick review, I would like to discuss the website by Lin and Don Donn, http://earlyhumans.mrdonn.org/evolution.html.
This website is part of a much larger website that is filled with a lot of information on all sorts of history. As Mr. Donn states, they do not claim to be experts in anything, so do not claim everything on the site is correct, although they do try. It is clear they have put a great deal of time and effort into making a substantial site with the honest intention of providing accurate and useful information to teachers. They have won awards for an impressive site. However, in the evolution of humans, they seriously fall down.
The early humans website has several links to good resources. Unfortunately, it has two things that destroy the science educational credibility of the site completely. The first is a link to a presentation teaching Biblical creationism, a subject that has no place in a public school as it is both scientifically invalid and pushes one specific religious view, which is illegal in the United States. Regardless of whether one believes in creationism or not, it is not legal to teach a specific religion in public schools and it is especially not valid to teach that religious view in a science class. The only way to make this legal would be to teach the creationism stories of every other religion equally, without comment as to which one the teacher believed, which would be impossible. Even then, it would have to be in a religious studies class, not a science class. If we are to preserve everyone’s First Amendment rights to freedom of religion, we simply cannot have government-run public schools teach one religious view and we certainly cannot teach that view as a scientifically valid theory. I am hitting this point especially hard because it is a serious point of controversy in the United States, but it should not be. Keeping creationism out of the schools is not an attempt to suppress anyone’s views. It is an attempt to preserve everyone’s right to make their own religious choices without government interference.
That leads me into the second problem, one which is stated boldly right up front. One of the big problems we have in science literacy is that many people do not understand the difference between the colloquial use of the term “theory” and the scientific meaning of the term. To quote the website: “A theory is a guess based on some facts. Remember a theory is not proven. One of the great controversies of our time has been the theory of evolution.” This is massively wrong in two areas.
The term they have defined is NOT a theory. What they defined was SPECULATION. Anyone can come up with an idea, but that does not make it a scientific theory. First, one must have a hypothesis, which is a testable idea, based on observation, that explains a relationship between two or more measurable things. There are two critical parts to this. The observations, so it must be an attempt to explain something we actually see in the real world. Second, that explanation must be testable. If there is no conceivable way to test it, the idea remains in the realm of speculation and can never be taken as a scientific theory, or even a valid hypothesis.

Great hypothesis, but not the way to do science.
Once one has a series of hypotheses that have been tested by many people, none of whom have been able to disprove the hypotheses, one can formulate a scientific theory. That theory ties the hypotheses together, explaining numerous detailed observations into an explanatory framework that applies broadly. An example of this is the Theory of Gravity. Numerous observations were made showing gravity exists, there is no doubt about that. Many observations showed precisely how it worked and the relationship of different masses to each other, both on earth and in the universe as a whole. However, to make a theory, we needed more than these observations, we needed a way to accurately describe and predict these relationships. Isaac Newton discovered a mathematical equation that could be used to predict the motions of the planets. That equation was then tested many times and found to be valid everywhere, at least at the speeds attained by most things in the universe. Einstein went further with his Theory of Relativity, which extended our understanding of gravity into realms beyond the experience of everyday existence. Even here, these started out as hypotheses, requiring many people to test over and over gain. Not only has no one been able to prove them wrong, but no one has come up with an explanation that better fits the data. Therein lies the key, testing and testing and basing the acceptance of the theory on data, evidence that either supports or disproves the theory. Without that, it is not a theory.

As such, there is no Law of Gravity. We know it exists, it is fact that is undeniable. The Theory of Gravity provides a framework in which gravity works that has been put to the test. In a similar fashion, there is no Law of Evolution. We know it exists, it is a fact that is undeniable. Why? Because the idea that biological life forms change over time is something that cannot be argued against. All one has to do is acknowledge we are not clones of our parents, or look at the diversity of changes brought about by dog and cat breeders, sheep and cow breeders. We see biological change all around us. Evolution is therefore a fact, just like gravity. The Theory of Evolution put forth by Darwin is more properly called the Theory of Natural Selection, which explained this change through the aforementioned natural selection. It has been tested numerous times and shown to work. Is natural selection the only way in which species change? No, but it is a major mechanism. But the point here is that it has been tested and retested. Like all scientific theories, it is not simply a guess based on a few facts. It is permissible to argue about specific mechanisms, but trying to argue whether or not evolution occurs is like arguing whether or not the earth is flat or that we need air to survive.

Mystery Monday Fossil
Today’s Mystery Monday fossil is related to some recent paleontology news that I am excited to talk about here because it has put Arkansas fossils on the map in a much bigger way than ever before. See if you can figure out what we have here.
Fossil Friday: We may have bats in the belfry, but we’re not crazy.
I know this is getting posted late, sorry about that. I got an unexpected request by a local news reporter to be interviewed about a fabulous new shark fossil found in the Ozark Mountains of Arkansas. I will have to write about that soon. But that’s for later. For today, we have something very different.
This week’s Mystery Monday fossil was a tough one. People are much more used to seeing them alive than as fossils. When people do see them, they usually don’t like them, which is a shame. These little guys are very misunderstood creatures and we owe them a debt of gratitude.
 This picture is from Arthur Harris of the UTEP Biodiversity Collections. It is a jaw from Eptesicus fuscus, the Big Brown Bat. Why a bat, you ask? Because April 17 was National Bat Appreciation Day. Most people do not appreciate bats, holding onto the myth that bats like to fly into people’s hair and give them rabies. In truth, bats do their very best to avoid people. If you weighed less than a pack of cigarettes with fragile bones, would you annoy something that outmassed you by at least 2000 times? The simple fact that bats are the only mammals that can fly is pretty cool, although perhaps less so when one considers that bats make up over 1000 of the approximately 5500 species of mammals. Nevertheless, being able to fly using hands that are each bigger than its body is appreciation-worthy. The fact that their wings are giant hands helps explain why they can fly with such aerobatic skill. We have mechanoreceptor’s in our skin called Meissner’s corpuscles, which allow us to have a well-developed sense of touch in our fingers. Bats have these in huge numbers all over their wings, using them to detect the air pressure on the wings as they fly. The wing is essentially a giant finger tip, sensory speaking. They also have tiny hairs that are very sensitive to airflow over the wings.
This picture is from Arthur Harris of the UTEP Biodiversity Collections. It is a jaw from Eptesicus fuscus, the Big Brown Bat. Why a bat, you ask? Because April 17 was National Bat Appreciation Day. Most people do not appreciate bats, holding onto the myth that bats like to fly into people’s hair and give them rabies. In truth, bats do their very best to avoid people. If you weighed less than a pack of cigarettes with fragile bones, would you annoy something that outmassed you by at least 2000 times? The simple fact that bats are the only mammals that can fly is pretty cool, although perhaps less so when one considers that bats make up over 1000 of the approximately 5500 species of mammals. Nevertheless, being able to fly using hands that are each bigger than its body is appreciation-worthy. The fact that their wings are giant hands helps explain why they can fly with such aerobatic skill. We have mechanoreceptor’s in our skin called Meissner’s corpuscles, which allow us to have a well-developed sense of touch in our fingers. Bats have these in huge numbers all over their wings, using them to detect the air pressure on the wings as they fly. The wing is essentially a giant finger tip, sensory speaking. They also have tiny hairs that are very sensitive to airflow over the wings.
Despite people’s antipathy towards them, bats are very valuable. Without them, we would be eaten alive by mosquitos and all manner of bugs. A little brown bat can eat up to 1000 mosquitos in one hour and a single colony of big brown bats can eat 33 million rootworms in one summer. Big brown bats eat all sorts of insects and are adept at catching them in mid-flight. The spiky, ridged teeth are an adaptation to crunching the hard, chitinous covering of beetles and such types.Bat guano has been used for numerous products, everything from fertilizer, gasohol, antibiotics, and gunpowder. The big brown bat in particular seems to thrive in areas with humans, often setting up colonies on man-made structures. So if you have bats in your belfry, they are probably this type. But don’t worry, they’re helping you.

cmns.umd.edu

Even though they are called big brown bats, they are only really big compared to the little brown bat. They attain body lengths up to 13 cm (5″), with a wingspan of a whole foot or more, which is large for a bat. Their fur ranges from tan to a dark chocolate color, with black skin, making their wings black, naturally. Basically, think of your average bat (although a bit smaller than most people’s hyperactive imaginations) and you will be thinking of Eptesicus. They have excellent hearing and can use echolocation to find their way around, although the idea that bats are blind is a complete myth, they see just fine. Because they are small and fly, they need to eat a lot. So to make up for an insatiable appetite, they sleep a lot during winter when food is scarce. A lot. Big brown bats can hibernate over 300 days, almost a whole year. Even so, they can tolerate cooler temperatures than most other bats and can come out of hibernation during winter if need be. They can live for almost 20 years, but most die within their first year. Winter is rough for bats, especially when they are small. They aren’t mature until their second or third year and only have typically only two offspring a year, so they do not have a high reproductive rate. Even if they have a low reproductive rate, their heart rate is staggering. They go from over 400 bpm at rest to well over 1000 bpm in flight. One wonders how they prevent their heart from exploding. It is often said that bats have legs with knees bending the opposite direction of other mammals. This is not really true. What they have are legs which are turned at the hip so far they look backwards.

Palaeochiropteryx, a fossil bat from the Messel Shales. Fossilmuseum.net.
Fossils of this animal can be found throughout the southern United States all the way down to Venezuala (Kurta and Baker 1990). The genus Eptesicus can also be found throughout Europe. No bat is very common in the fossil record, but these are the most common and widespread. They also seem to be expanding their range. The fossils are found throughout Pleistocene deposits for the past few million years. The current range goes all the way up into Alaska, from the Pacific to Atlantic coasts,and throughout the Bahamas and northern South America. if you want to find fossil bats in Arkansas, I recommend you go into the caves in the Ozark Plateau. Any place you can find modern bats living, there is a decent chance that if you dig down into the floor of the cave, you will find bat fossils.

One down, only 33 million more to go this year. http://www.life.illinois.edu/paige/bats.html
Kurta, A., R. Baker. 26 April 1990. Eptesicus fuscus. Mammalian Species: No. 356, pp.1-10.
Mystery Monday, Tax week edition

For today’s Mystery Monday fossil, we are looking at an animal who is being celebrated this week. No, it’s not a rabbit, Easter is next week. For scale, the bar on top is marked in millimeters, so we are looking at a wee little creature. If getting your taxes done is driving you crazy, spend some nice relaxing time with a fossil.
Fossil Friday
On Monday the fossil shown below was put up. How did you do? We got some very good suggestions about it possibly being a horn or a tooth. It isn’t a horn, not exactly. It does also bear a remarkable resemblance to some spinosaur teeth I’ve seen, which was one of the guesses. But it’s not a tooth either. Congratulations to Laurenwritesscience, who really knows her fossils for correctly identifying it as a horn coral.
 Modern day coral are pretty much all in the group Scleractinia, meaning stony rays, known as the stony corals because they make an external skeleton out of aragonite, a form of calcium carbonate, very similar to calcite. They are the major reef builders in modern oceans. But it wasn’t always that way. Before scleractinians, there were tabulate corals and rugose, or wrinkly corals. Rugose corals, like modern corals formed both large colonies and solitary structures. The solitary rugose corals, like this one, are called horn corals, obviously for their commonly stated similarity to horns.
Modern day coral are pretty much all in the group Scleractinia, meaning stony rays, known as the stony corals because they make an external skeleton out of aragonite, a form of calcium carbonate, very similar to calcite. They are the major reef builders in modern oceans. But it wasn’t always that way. Before scleractinians, there were tabulate corals and rugose, or wrinkly corals. Rugose corals, like modern corals formed both large colonies and solitary structures. The solitary rugose corals, like this one, are called horn corals, obviously for their commonly stated similarity to horns.
Rugose corals first appeared in the Ordovician Period roughly 47o million years ago, give or take 20 million years. They became important reef builders in the Devonian Period and remained a highly visible presence until the end of the Permian about 250 million years ago. They, like almost everything else, succumbed during the Great Dying, as the mass extinction of that time is called. As a result, if you want to find them in Arkansas, you should try the marine limestones in northern Arkansas, throughout the Ozarks. The best place to find them are the Mississippian rocks, such as the St. Joe, Boone, and Pitkin Formations.

Life cycle of a rugose coral. Picture from the Kansas Geological Survey
Rugose corals, like all corals, are more than just the stony skeleton most people think of when they see coral. Inside, the animals themselves grow as polyps sitting, in the case of horn corals, a stony cup called a calyx, which they constantly build up as they grow. When alive, the pointy end went down, permanently attached to the surface upon which it grew. Inside the calyx, they grow septae, little walls used to provide structural support for the skeleton and to hang themselves on (while this may cause one to picture a very depressed polyp, a better visual image would be hanging pictures or cabinets on a wall). Rugose corals typically grew six main septae, forming six compartments. Four of the compartments were further subdivided by minor septae. All of these septae radiated from a central axis, or rod supporting the whole structure. The four main compartments is one reason why they are sometimes called Tetracorallia.

Growth of septae in rugose and hexacoral. Paleo.cortland.edu.
This is one of the key characteristics that separate them from modern day coral, which are members of a group called Hexacorallia, noted for having six main septae and six main compartments, all of which are furhter subdivided with minor septae, thus having six-fold symmetry (you can draw six lines through the center of the organism in which one side is a mirror image of the other). Hexacorallia also include the sea anemones, also noted for having six lines of symmetry. The Rugosa, on the other hand, due to the four compartments, often exhibited only bilateral symmetry, meaning there was only one plane you could draw to find a mirror image.

Coral polyp anatomy. Fossilmuseum.net.
The polyp forming the living organism was a tiny creature with a simple sac-like digestive tract, meaning there is only one opening. Food passes into the mouth and waste comes right back out the same way. This is the typical pattern seen in almost all of the Cnidarians, the group in which corals are a part, along with animals like the jellyfish. Like the jellyfish, coral polyps have nematocysts, stinging cells they use to capture prey. Surrounding the mouth, and where one would find the nematocysts, are a ring of tentacles, forming a flower appearance, the reason the group containing all corals is called Anthozoa, the flower animals. A lot of group names and you might be getting lost, so I’ll summarize the names: Horn corals are a part of the Rugosa, or rugose corals, also called Tetracorallia, which are a part of Anthozoa, along with all the other corals and sea anemones, which are a part of Cnidaria, which also includes the jellyfish among others. The modern corals are mostly scleractinians, or stony corals, which are a part of Hexacorallia, which are Anthozoans and thus Cnidarians (which you may also see referred to as coelenterates). Like all members of the Anthozoa, rugose corals almost assuredly reproduced asexually by budding or sexually by releasing both sperm and eggs. In many corals, most of the polyps in an area will do this at or near the same time, called broadcast spawning, creating a massive cloud of eggs and sperm floating around the reef. I have read this is a fascinating sight, if rather disturbing to some. Why do they do this? Because everything else in the reef considers them good eating, so the only way to ensure that some escape long enough to survive and grow is to simply overwhelm them with sheer numbers. This idea is often called predator satiation. It also gets around the fact that coral can’t move, they are the ultimate homebodies. The only hope they have to meet a good mate is to just put the seeds of their offspring into the wind, or in this case, currents, and hope for the best.
Most corals do not depend just on what they can capture or filter out from the water. They also have a mutualistic relationship with photosynthetic dinoflagelates and zooanthellae, or single celled algae. They algae get a safe place to live and nutrients, while the coral gets the products of the photosynthesis, such as oxygen, sugars and amino acids. Normally the coral don’t depend upon the extra energy derived from the photosynthesis, but when times are tough, the added supply can see them through, allowing both the coral and the algae to survive longer. The mutualism certainly provides a much more abundant resource supply for the coral, allowing them to grow more efficiently. We don’t know if rugose coral did this, but it is reasonable to assume they did for now. Of course, this means that corals who use this relationship are restricted to shallow, clear waters conducive to the growth of algae. We do know that they formed close relationships with stromatoporoid sponges. The sponges formed the backbone of the reefs, with the coral growing within the sponge reefs. What sort of association beyond that is not known though.

Life in the Devonian seas. UMich Mus. Natural History.
Mystery Monday

Time for another Mystery Monday fossil! See if you can figure out this wrinkly rock.
Fossil Friday, Having a Blast
It’s Friday, which, along with the anticipations of the weekend, means it’s time for the reveal of this week’s Mystery Monday fossil. We’ve had guesses of starfish, aka sea star, and sea urchins. Both are close. Were you able to guess it?

This fossil is of Pentremites, a echinoderm in the group Blastoidea, so a relative of both sea stars and sea urchins. Like them, you would have found them in shallow marine communities in fairly clear water, if they still lived. Blastoids are what you might get if you crossed a crinoid (another echinoderm) with a sea urchin, but we’ll get to that. This particular image was taken by Dr. Richard Pasilk, of the Humboldt State University Natural History Museum. You can find it and many more fascinating images at the Paleoportal.org fossil galleries.
Echinoderms, or “spiny skin,” have been referred to as walking castles because most of them form plates and spines of calcium carbonate that lock together, forming a mobile fortress. Echinoderms include starfish, sea urchins, crinoids, and holothuroids, or sea cucumbers. Echinoderms are known for having tube feet, a part of their water vascular system. If anyone has seen hydraulic mechanical systems, you know how these work, by pumping water in and out of tubes to change the water pressure, allowing the tubes to extend or contract. They don’t have much in the way of nervous or sensory systems, although sea stars do have rudimentary eyes allowing them to see, albeit very poorly. At least some sea stars can turn their stomachs inside out to eat, and sea cucumbers can basically eviscerate themselves, ejecting their guts through their anus, to gross out potential predators. Sea cucumber poop is also very important for coral reefs, so be a hero, save the sea cucumber, save the ocean.
Blastoids grew on long stalks like crinoids formed of many flat disks, but instead of having fairly disordered plates that formed a rough ball-shaped shell called a theca, the plates forming the blastoid thecae were nicely ordered, arranged in a shell that many have thought resembled a hickory nut. This ordered, integrated theca is much more similar to the echinoid sea urchins than it is the crinoids. It has the advantage for fossil hunters that it held together better, meaning that they are much easier to find than crinoid thecae, which pretty much scattered across the sea floor as unidentifiable calcite crystals as soon as the animal died, unless they were killed by being buried.

Reconstruction of Asteroblastus volborthi by Ernst Haeckel, 1899-1904.
The mouth is located at the top, surrounded by five grooves called ambulacra. Coming off the ambulacra were a series of feathery appendages called brachioles, which would filter particles from the water, much like the feathery arms of the crinoids. Between the start of each ambulacra sat an opening. Four of them led to the respiratory system, consisting of complexly folded structures called hydrospires. Loosely fold a piece of paper a couple of times, then roll it up and you will get an idea what it looked like. Water would flow from the brachioles into your paper hydrospire between the edges of the paper and out the top of the tube. The other one was the anus, so the digestive system was U-shaped, with the mouth and anus adjacent to each other.

The fossil record of echinoderms is extensive, starting in the Cambrian over 540 million years and possibly as far back as the Ediacaran around 600 million years ago. The fossil record of the blastoids is somewhat debated. Whereas some sources say they originated in the Ordovician, most put the oldest confirmed blastoid in the Silurian, roughly 425 million years ago. They became abundant in the Mississippian Period and were persistent members of a diverse shallow marine community until they died out by the end of the Permian Period a little over 250 million years ago, along with most of the world in “The Great Dying.” In Arkansas, as in the rest of North America, blastoids were common and diverse in the Mississippian Period, also known as the Lower Carboniferous Period, although they became rare in the Pennsylvanian, the Upper Carboniferous Period. Arkansas has some of the only Pennsylvanian blastoids in North America.

Look for rocks like this. Pitkin Limestone along Hway 65. http://www.geology.ar.gov
If you want to look for them in Arkansas, the best places to go would be the Mississippian age limestones in the Ozark Plateau, such as the Pitkin Limestone and the Boone Formation, and the early Pennsylvanian age limestones, such as the Brentwood Limestone of the Bloyd Formation. Follow Highway 65 north towards Leslie and Marshall and stop at any roadcut through the Ozarks showing whitish rocks and you stand a decent chance of finding them. Just don’t collect in the National Forests and watch the traffic.