Home » Posts tagged 'dinosaur' (Page 2)
Tag Archives: dinosaur
Day 1 at SVP
 I attended the annual meeting of the Society for Vertebrate Paleontology (SVP) last week. This is the yearly meeting where those studying anything with a backbone, from fish to furballs get together to talk about what they have been doing and see what others have found (although admittedly, most of the press goes to dinosaurs). Over the next few posts, I will share brief snippets of what I learned. If you read anything that sounds particularly interesting to you and would like me to discuss it in more detail, just let me know in the comments section and I can expound on the topic.
I attended the annual meeting of the Society for Vertebrate Paleontology (SVP) last week. This is the yearly meeting where those studying anything with a backbone, from fish to furballs get together to talk about what they have been doing and see what others have found (although admittedly, most of the press goes to dinosaurs). Over the next few posts, I will share brief snippets of what I learned. If you read anything that sounds particularly interesting to you and would like me to discuss it in more detail, just let me know in the comments section and I can expound on the topic.
So what does a scientific meeting like this look like? There is always a dealers’ room, where you can find an array of book publishers, such as Indiana Press and CRC Press, and supply companies, such as Paleo-Tools and Bone Clones, and many others all hawking their wares. You will meet artists such as Luis Rey, learn about what’s going on at different publications, such as PLOS One, and upcoming meetings. There are the business meetings, social events, and award banquets. There are field trips to local paleontolgical sites (this meeting was in Los Angeles, so the La Brea tar pits and the Los Angeles Natural History Museum were highlights). There is also the chance to converse with paleontologists from all over the world, a venue wherein colleagues can talk face-to-face and forge new connections, where students can get their feet wet presenting at an international conference and talk to students of other professors to learn about potential graduate and post-graduate opportunities, which professors make good mentors and which ones to avoid. And of course, there are the talks and poster sessions, where you can hear about the research people are doing right now. If you want to know the current state of the field, this is the place to go. Every day for four days, three sessions run concurrently, each covering 16 talks a day, plus over 120 posters are presented every day. By the end of which, if you haven’t found enough new information to send your brain into overload, you simply haven’t been trying. This is also a relatively small meeting, with only 1500 or so attendees, unlike some scientific societies which have meeting attendances over 20,000.

Bosch Institute Neuroscience meeting, 2007. This is a bad design for meetings. Please, just stop.
Before I get into a short recap of the talks I attended (there were many more I would have liked to attend, but I have not yet perfected cloning myself), I will digress a moment for a quick comment about the meeting room setup. My first thought upon seeing the room arrangement was Happy day! All the meeting rooms are right next to each other; not on a different floor, on the other side of a giant building, in another building, or in another facility entirely, unlike some other meetings I’ve attended. This makes popping from one session to another to see different talks much easier. The downside to this is that hundreds of other people all have the same idea and are trying to go through the same hallway, trying to get around the other attendees who are chatting with friends and colleagues. For those of you that are accustomed to New York subways, this is no big deal, but for some of us, it can be a bit claustrophic as personal space shrinks to microscopic proportions. The other rant I have is that, after all this time, they still have not learned how to lay out a room for the talks. They insist on preparing the rooms as though everyone will calmly file in before the talks and sit there until the break, providing long rooms with the speaker at one end and two long columns of interlocking chairs set in rows up to 20 seats long. however, this is a scientific meeting with many short talks. people stream in and out constantly. So what happens is that all the edge seats are quickly taken and great, yawning chasms of emptiness are left in the center. Why no one ever thinks to place the speakers in the middle of a long wall, with many columns of seats with no more than 6-7 seats to a row, allowing ample space for people to move, is beyond me. They might not be able to put as many seats in, but that hardly matters if no one can get to the seats in the first place. Now on to the talks!

Sauropod nest, by Mike Ellison
In the first session, all the talks dealt with the problems of ontogeny in interpreting the fossil record. Ontogeny, how an animal grows from fertilized egg to old age, can cause several problems in the fossil record. When all you can see is a fossil, it can be very hard to tell if you are looking at an adult or juvenile. Many animals can change so much during development that the juveniles can look like completely different species. Hans Larsson presented about a possible way to figure out rough ages of an animal by looking at how different bones in the skull covaried, or how they changed shape in relation to each other as they grew. Jack Horner warned against assuming an animal is full-grown unless you have solid evidence of that from the bones across the skeleton and Mark Goodwin showed how difficult that can be with pachycephalosaurs, but Holly Woodward found that at least some dinosaurs reached sexual maturity long before they reached “adult” size and in fact, never stopped growing, so it is important to keep in mind just how one defines “adult.” David Evans showed the problems of using juveniles and adults in the same phylogenetic analysis, in that the relationships of species identified using juveniles were very ambiguous and unstable, often showing up as more ancestral than an adult of the same species, sometimes substantially so. John Scannella found that using just one juvenile specimen of Triceratops in a phylogenetic analysis resulted in that specimen not being identified as a ceratopsian at all, but if a sufficient number of juvenile specimens of different ages were included, they all fell out together in their expected relationship. This indicated the ontogeny problem can be dealt with if you have enough samples, as well as indicating what characters are newly evolved versus being evolutionarily stable. Zachary Morris extended this to show that not all individuals grow up the same way, there was no set pattern of development that all individuals within the same species followed to adulthood, so trying to order specimens into an ontogenetic sequence requires a large sample size. Also, contrary to an earlier research that got a lot of press, Torosaurus is probably not the same species as Triceratops. Robert Reisz showed very fast growth in prosauropod embryos and P. Martin Sander estimated that sauropods could grow to sexual maturity in 16 years, attaining 90% of their maximum size within 32 years. This comes out to an average growth rate of 4 kg/day, which is similar to modern large mammalian herbivores, but he cautioned this was likely overestimating the ages and underestimating the growth rates. Ken Dial discussed the role of predation in the development of flight, with evidence indicating that if predation pressure was relaxed and there was sufficient food resources, birds commonly became flightless.He made the interesting point that everything is a transitional environment; that to truly understand an organism,one has to examine all aspects of their habitat and you can’t afford to get to narrowly focused. Denver Fowler found that animals occupying different niches at different ages were more susceptible to extinction. Caroline Stromberg found that, contrary to popular belief, the development of grasslands didn’t actually correlate very well with high-crowned, hypsodonty teeth.
Wikipedia.org
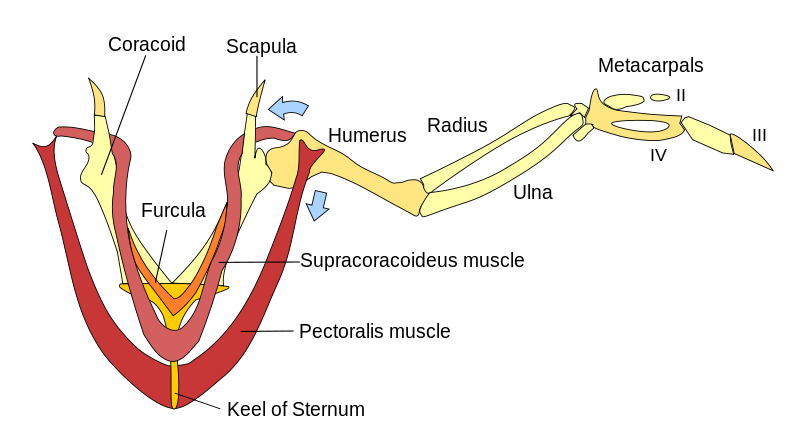
Sandy Kawano looked at how locomotion changed between fin and feet and found that the total forces between front and hind limbs was fairly constant, but acceleration came from the pectoral fins at first, but as the hind limbs developed, more of that motive force came from the hind legs, with the front legs taking on more of a regulatory role guiding maneuverability. Karen Sears found that limb evolution became more modular, in that different parts of the limb evolved at different rates and times, so that each part could be considered as evolving relatively independently, with higher levels of evolution near the ends of the limbs. Paul Sereno discussed the evolution of the coracoid bone in the shoulder and how its loss in almost all mammals created a more flexible and faster, but weaker joint. He opined this is why bats don’t grow to the size of birds or pterosaurs and that it allowed dinosaurs to get bigger and carry more weight than mammals.
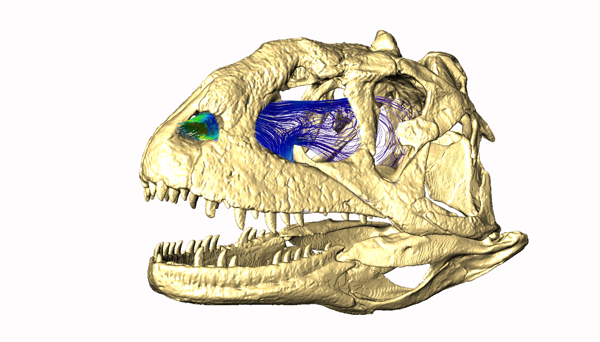
Model of air flow through the nasal passages of Majungasaurus. Image and work by Jason Bourke
Ali Nabavivadeh reported on his studies on the evolution of jaws in ornithischian dinosaurs, finding they talked the problem in a variety of different ways. Lucas Spencer noticed that all early ornithischians were in Gondwana and that dispersal explains more of their biogeography than vicariance (speciation via the emergence of physical barriers). Mark Loewen reported on the biogeography and phylogeny of ankylosaurs, while Victoria Arbour discussed the evolution of the ankylosaur tail club. Philip Currie reported on the smallest known articulated ceratopsid fossil, while Andrew Farke reported on ceratopsian biogeography. Thomas Carr found evidence to support anagenesis (direct ancestor-descendant relationships) in some tyrannosaurs, but not all, while Brandon Hedrick found that fossilization processes made one species of psittacosaurs look like three different ones. Jordan Mallon found little evidence in pachycephalosaur skulls to indicate they lived in upland areas, as is often stated, and Jason Bourke did some very nice computer modeling showing how turbinates (structures within the nasal passages often associated with endothermy, aka warm-bloodedness) work to channel air through nasal passages,using that to find support for respiratory turbinates in pachycephalosaurs.
This all only recounts brief snippets of the talks I attended on the first day and doesn’t include the many posters I looked at. I will cover Day 2 in my next post. If you want more information on any of these snippets, leave a comment and I will be happy to expound upon them.
Forum Friday, winging our way to LA
Paleoaerie is off to Los Angeles next week to attend the annual meeting of the Society for Vertebrate Paleontology. If it has a backbone, it will be discussed there (if it is still alive, the discussion will be linked to fossil relatives, but I didn’t want to unnecessarily limit it, because I’ve seen gators, chickens, dogs, sharks, and everything in between discussed there in relation to their fossil ancestors; I’ve even heard and presented on bacteria, insofar as they help make fossils).
As a result, I’ve only done one blog post on Paleoaerie since last Forum Friday and I won’t be able to post anything other than maybe comments about the meeting next week, so I wanted to get a forum post out before I go, although I realize that some of you reading this may not see it before Monday, what with the lateness of the hour this got posted.
So without further ado, here we go. On Paleoaerie, we talked about the book, Scaly spotted feathered frilled: how do we know what dinosaurs really looked like?” by Catherine Thimmesh, a great book for any budding paleoartist you know.
Over on Facebook, we celebrated Member Night at Mid-America Museum and their new dinosaur exhibit, Reptile Awareness Day, and Geologic Map Day. We learned about the Backfire Effect and why telling people facts is not always convincing and ways to frame your arguments that may work better. We warned against the ResponsiveEd curriculum, as well as Stephen Meyer’s book, Darwin’s Doubt, with some interesting new studies on the preCambrian life leading up to the Cambrian explosion.
We learned more about Tyrannosaurus rex and what we still have to learn, and we met Joe the dinosaur, the most open-access dinosaur ever. We learned how rapid evolution in one organism can cause a cascade of reactions throughout the ecosystem.
 We saw a whole host of dinosaurs in 3D, as well as horses and a lot of cool videos from the Science Studio. We saw a new animation explaining how the evolution of life affected the early atmosphere, oceans, and which rocks were formed.
We saw a whole host of dinosaurs in 3D, as well as horses and a lot of cool videos from the Science Studio. We saw a new animation explaining how the evolution of life affected the early atmosphere, oceans, and which rocks were formed.
We learned about the usefulness of evolution in medicine, how allergies can save your life, and that sharks, contrary to popular opinion, suffer from cancer just like the rest of us. We also saw a robot made completely out of prosthetics made for humans. Where will we go from here and how much farther can we go?
May your Halloween be filled with spooky fun!
Myths and Misconceptions #1: Allosaurus rex, or That is NOT a Dinosaur
When I worked at the Denver Museum of Nature and Science as a volunteer in the educations collection (if you ever get the chance to volunteer at a museum, do it, it was a lot of fun and very educational), I happened upon a man with a young boy by his side, whom I presume was his son, looking at an impressive skeleton. I thought how great it was the man took time to bring his son to the museum. But when the boy asked what the skeleton was and the man answered, “Allosaurus rex,” my opinion of the experience dropped. I’m still glad he brought the boy to the museum, but it could have been so much better.

Allosaurus rex? Actually, a blue whale. Definitely NOT a dinosaur. Also, not DMNS. Really, the London Natural History Museum. Photo by author. Click to enlarge.
So what was wrong with what he said? There is no such creature as an “Allosaurus rex.” There is an Allosaurus and there is a Tyrannosaurus rex, but not the two together. It also did not help that the man was standing directly in front of the plaque that read, “Fin whale.”
Much has been written on what a dinosaur is and what is not, but considering the extreme levels of confusion in the general populace, I thought it worth discussing it here. Not everything that is a big skeleton and/or extinct is a dinosaur. In addition to the above whale, I have heard dimetrodons, pterosaurs, saber-toothed cats, mammoths, giant rhinos, mosasaurs, modern elephant skeletons and many other things called dinosaurs, when in reality, NONE of the previous creatures qualify as dinosaurs.
So, if none of those are dinosaurs, what is? What are some of the things people think about when they try to define dinosaurs? The first thing that most people think about is a giant, scaly reptile. However, some dinosaurs were the size of chickens, so not all of them were big. Many also had feathers, so the scaly motif is not altogether correct either. They are reptiles, but unlike anything most people would consider a reptile today.

Quetzalcoatlus northropi. UT Museum. Not a dinosaur either. Photo by author.
Other traits people often use to distinguish dinosaurs are that none of them were aquatic and none of them flew. These aren’t really true either. Some will say they only lived in the Mesozoic Era and died out at the end of the Cretaceous Period 65 million years ago. Again, false. It is true that most animals considered dinosaurs did indeed live only in the Mesozoic Era, but not all died out at the famous K-T extinction event. Some of you may be thinking to yourselves the author has no idea what he is talking about. Nevertheless, as we will see, some dinosaurs are quite at home in both air and water, especially the ones that lived past the Mesozoic.
Even if all these things were true about dinosaurs, none of it really matters. These traits are all distinguishing characteristics that are generally true about most dinosaurs, but not necessarily true about all. What defines an organism is not the same as the diagnosis. Definitions are done by evolutionary relationships. Diagnoses tell us how we can recognize them, what distinguishing characteristics can be used. However, distinguishing characteristics do not necessarily describe all the members of the group. For instance, one might distinguish all members of a family by their last name, but if people get married and change their last name, they do not suddenly stop being a member of the family. A daughter, for example, is defined by who her parents are, not by what name she has. Likewise, her distinguishing characteristics tell us who she is as an individual and can give us clues to her family relationships, but her definition as a daughter depends solely on her relationship to her parents.
The discredited and completely unaccepted haemothermia hypothesis: Mammals and birds are both warm-blooded; therefore, they must be related. I.e., superficial similarities can be misleading. Image from wikipedia.
I should note here that it wasn’t always this way. Back in the old days, organisms were grouped by similarity, who looked like whom, so at that time, there was really no difference between definition and diagnosis. But as people studied organisms more closely, they found that a lot of times, superficial similarities masked deeper differences, which indicated they weren’t really related at all. So for most of the last century, scientists have tried to find real, evolutionarily related groupings to build something akin to a geneology of life. To do so, it became evident that definitions and diagnoses of groups had to be different, so for the last couple of decades, definitions have been based on relationships and diagnoses based on characteristics. This system of classification by defining groups according to their relationships is called phylogenetic systematics.
So, how then do we define a dinosaur? The standard modern definition of a dinosaur has been stated as the most recent common ancestor of Triceratops and modern birds and all of its descendants (Padian, 1997). Note this means that all birds are, by definition, dinosaurs. So it is obvious that some dinosaurs are quite small (e.g. hummingbirds), flew (most birds), and could reasonably be called aquatic (e.g. penguins). The reason for this is because the earliest birds are clearly closely related to animals, such as Velociraptor and its close relatives, which are indisputably dinosaurs. If one took a picture of Archaeopteryx which had no feathers to a bunch of elementary school kids and asked them to identify it, the general answer is that it is obviously a dinosaur, but put feathers on it and they call it a bird. They are thus both correct answers. Dr. Thomas Holtz has proposed a slightly different definition: the most recent common ancestor of Megalosaurus and Iguanodon and all of its descendants. Both definitions encompass the same groups of animals as far as our understanding goes, but Holtz’s definition includes the dinosaurs that were first discovered, so is considered by many to be a superior definition.
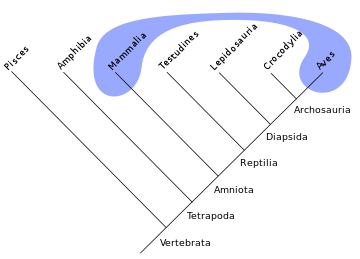
Dinosaur family tree, by Nobu Tamura (paleocritti.com). Click to enlarge. Note on trees like this, the branches can be pivoted on their branch points without changing relationships (e.g.,it makes no difference whether you say “your brother and sister” or “your sister and brother”), so anything in Saurischia is equally related to anything in Ornithischia. Thus, it doesn’t really matter what animal in each group you choose as the representative of each group as long as the groups as a whole remain intact.
Unfortunately, scientists don’t always talk about these terms correctly either, making the whole process confusing. Michael Benton, a leading paleontology researcher with numerous well-respected publications, got it wrong when he defined dinosaurs according to synapomorphies in The Complete Dinosaur. While synapomorphies are used in developing modern classifications, they are used for diagnoses, not definitions of groups, which Dr. Benton assuredly knows (as evidenced by the fact he normally states it correctly in his other works), but when someone is as prolific as Dr. Benton, the occasional error is bound to slip in now and then.
Synapomorphies are very important in figuring out relationships. Unlike any old similarity, synapomorphies are shared, derived characteristics, meaning that the character is the same in the organisms being compared because they share a common ancestor, i.e. they are derived from the same source. Of course, because it requires knowledge of the common ancestor, synapomorphies can only be identified AFTER one has a hypothetical relationship. Thus, to discover strong relationships, scientists will create (mathematically, using a computer because it is way too complex to try doing by hand for anything beyond a few species), all the possible relationships between all the organisms under consideration and then map all the characteristics they can onto every possibility. The relationships requiring the fewest inconsistencies and providing the simplest explanation is considered the most likely. Inconsistencies can occur due to convergent evolution (organisms not closely related developing similar characteristics due to similarities in environmental constraints) or homoplasies (characteristics changing back to a previous form), but with examination of enough characteristics, good relationships usually appear. Of course, the more data you can put into the analysis, the better the results normally get, which is why we continue to study and try to find new fossils and collect more data (and who wouldn’t want more fossils?).
This is all well and good, but how then do we recognize a dinosaur when we see one? Dr. Benton provided a good list of skeletal characteristics, which really won’t mean a lot to people who are not very familiar with skeletons and scientific terminology (but a basic description can be found here and Wikipedia has a surprisingly in depth description). Nevertheless, there are some generalities we can make. If we exclude birds, we can say the non-avian dinosaurs (that we know of) died out at the end of the Cretaceous Period. They all carried their legs underneath their bodies like mammals (and birds of course) and were decidedly unlike other reptiles. None were fliers, although the most bird-like ones got close, and none were aquatic, although some did indeed at least go wading. But remember, these are generalities and if we find a dinosaur with flippers, as long as it otherwise appears related to other known dinosaurs, it will still be a dinosaur. But if it does not fall within the relationships defined above, it will not be a dinosaur, no matter how much it may look like one.
I don’t have an online source for the Padian article, so here is the reference.
Padian, K. 1997. Dinosauria: Definition. pp. 175-179. In Currie, P.J. and K. Padian (eds.) Encyclopedia of Dinosaurs. Academic Press.
“Arkansaurus,” the only Arkansas Dinosaur

Welcome to the first of a series on Arkansas fossils. Arkansas is not generally known as a mecca for dinosaur lovers. Most of the dinosaurs in Arkansas are statues created by a man named Leo Cate, which have all the accuracy of the old plastic toys on which he based the statues, which is to say, not much (of course, he made them for enjoyment, not as anatomical models, so they serve their purpose). Nevertheless, dinosaurs are the first thing I get asked about when I give talks in schools, so I decided to start off with a discussion of our one and only dinosaur, called “Arkansaurus fridayi.”
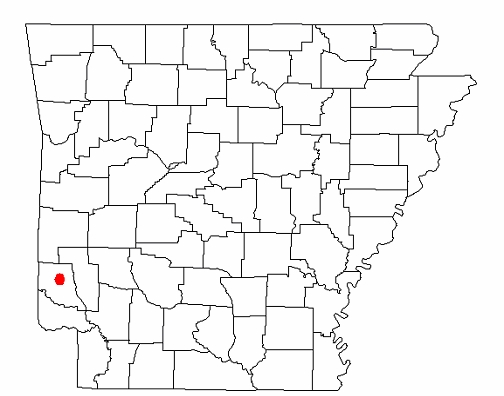 Ordinarily, I would not delve into how a fossil was found here, but because Arkansaurus is unique and illustrative of how many fossils are brought to the attention of science, a brief synopsis of the story of how it was brought to the attention of science may be of interest. In August, 1972, Joe Friday was searching for a lost cow on his property near Lockesburg in Sevier County, when he found some bones eroding out of a shallow gravel pit. He showed them to a Mr. Zachry, whose son, Doy, happened to be a student at the University of Arkansas in Fayetteville. Doy showed the bones to Dr. James H. Quinn, a professor at UA, who identified them as part of the foot of a theropod dinosaur. He contacted the Arkansas Geological Survey and Dr. Quinn, Ben Clardy of the AGS, and Mr. Zachry went back to the site where they found the rest of the bones. Dr. Quinn presented the bones at a meeting of the Society of Vertebrate Paleontology where he discussed the bones with Dr. Edwin Colbert, a noted paleontologist who was an expert in dinosaurs and vertebrate evolution. They came to the conclusion that the bones probably came from some type of ornithomimid, a group of ostrich-like dinosaurs (the name literally means bird-mimic), one of which, named Gallimimus, was made famous in Jurassic Park. Despite further excavations, no additional bones have been found. Dr. Quinn never officially described the bones, publishing only an abstract for a regional meeting of the Geological Society of America in 1973. It remained for Rebecca Hunt-Foster, now a paleontologist for the Bureau of Land Management, to publish the official description 30 years later in the Proceedings Journal of the 2003 Arkansas Undergraduate Research Conference.
Ordinarily, I would not delve into how a fossil was found here, but because Arkansaurus is unique and illustrative of how many fossils are brought to the attention of science, a brief synopsis of the story of how it was brought to the attention of science may be of interest. In August, 1972, Joe Friday was searching for a lost cow on his property near Lockesburg in Sevier County, when he found some bones eroding out of a shallow gravel pit. He showed them to a Mr. Zachry, whose son, Doy, happened to be a student at the University of Arkansas in Fayetteville. Doy showed the bones to Dr. James H. Quinn, a professor at UA, who identified them as part of the foot of a theropod dinosaur. He contacted the Arkansas Geological Survey and Dr. Quinn, Ben Clardy of the AGS, and Mr. Zachry went back to the site where they found the rest of the bones. Dr. Quinn presented the bones at a meeting of the Society of Vertebrate Paleontology where he discussed the bones with Dr. Edwin Colbert, a noted paleontologist who was an expert in dinosaurs and vertebrate evolution. They came to the conclusion that the bones probably came from some type of ornithomimid, a group of ostrich-like dinosaurs (the name literally means bird-mimic), one of which, named Gallimimus, was made famous in Jurassic Park. Despite further excavations, no additional bones have been found. Dr. Quinn never officially described the bones, publishing only an abstract for a regional meeting of the Geological Society of America in 1973. It remained for Rebecca Hunt-Foster, now a paleontologist for the Bureau of Land Management, to publish the official description 30 years later in the Proceedings Journal of the 2003 Arkansas Undergraduate Research Conference.
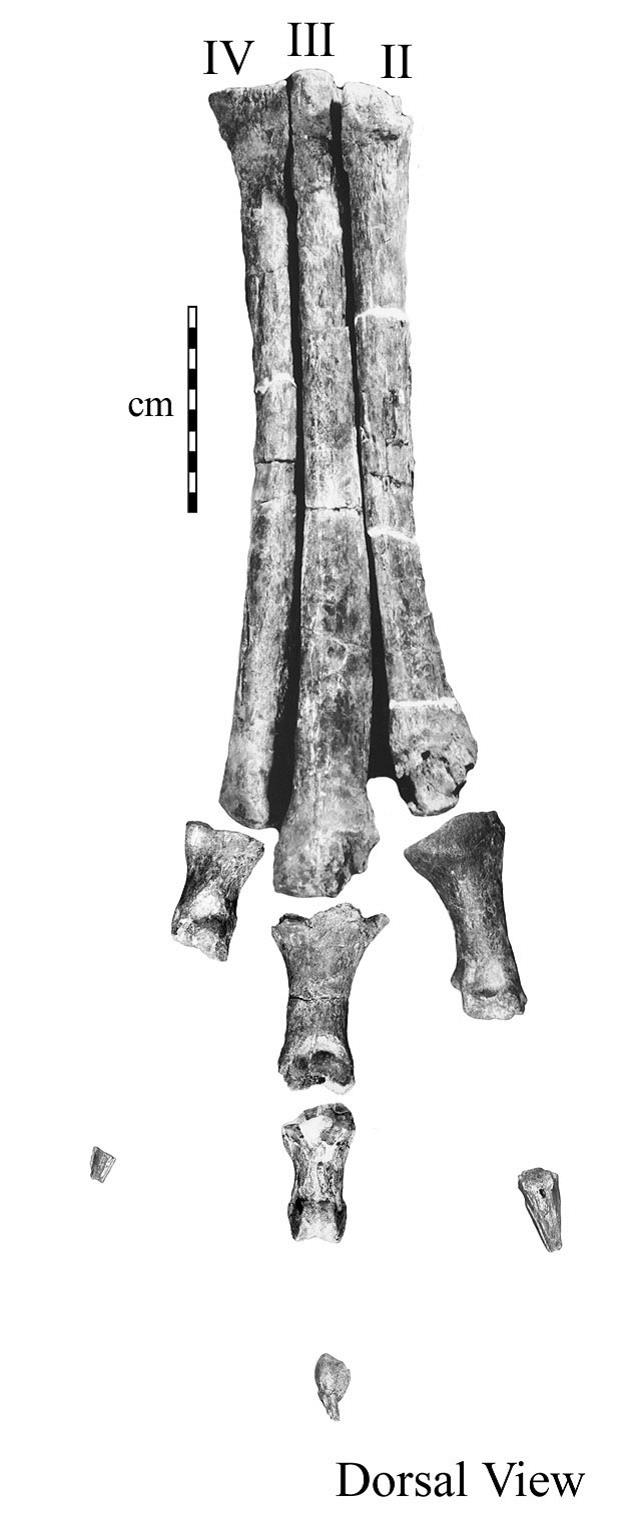

The first thing to know about this particular dinosaur is that “Arkansaurus fridayi” is not its real name. In fact, it doesn’t even have an official name. The reason for this is because all we have of it is part of one foot. Specifically, we have the metatarsals, a few phalangeal bones, and the unguals. In non-science speak, on humans, they would refer to the bones making up the front half of your foot. The metatarsals are the long bones the toes are attached to forming the front part of the arch, the phalanges are the toes, and the unguals are the bony cores of the claws. The pictures show the actual bones and a cast, in which the missing phalangeal bones have been restored. The real fossil has all the phalangeal bones connecting to the metatarsals and all the unguals, but a couple of the middle phalanges are missing. We have no ankle bones and nothing at all of the rest of the animal. With such little to go on, it has been difficult to determine exactly what kind of dinosaur it is, so no scientist has been comfortable giving it an official name yet. To add to the complications, not a whole lot of feet from theropod dinosaurs are known, so good comparison material is limited, and little is known about theropods in the southern United States to begin with. (Aside: dinosaurs are separated into two groups. The Ornithischia, which are comprised of the herbivorous, mostly four-footed dinosaurs; and the Saurischia, which include the giant, long-necked sauropods and the bipedal, mostly carnivorous theropods.)
So why only one foot? What happened to the rest of it? I’ll let Rebecca Hunt-Foster explain it, as she did an excellent job: “There are several possibilities that would explain the occurrence of a single foot at the Friday site. It is a possibility that the rest of the Friday specimen could be gravel on highway 24. Road crews could have cut into the Trinity Group (Ed. Note. The rock formation in which the bones were found) when excavating the Quaternary gravel that lies directly above it, when building the road in 1954. As another theory, the animal may have begun to decompose before its body was carried by water to the site of deposition. Consequentlly, bits and pieces could have been scavenged by predators in the Lower Cretaceous, resulting in only a single foot remaining for preservation. Finally, it is possible that the entire specimen was preserved but that most of the skeleton was lost to Pleistocene erosion.” So just think about that the next time you go driving down the road. What fossils might you be driving upon?
 Even if we don’t know for sure what it is, we do have some clues and can narrow down, at least a little, what it might be. What we know for sure is that it is some kind of coelurosaur. That, unfortunately, doesn’t help us a lot because coelurosaurs cover everything from little compsognathids to giant tyrannosaurs to modern birds, known principally for having bigger brains than earlier theropods, slender feet with three toes, and many of them had feathers. It does tell us it is not closely related to dinosaurs like allosaurs and spinosaurs, nor to early theropods like ceratosaurs and Coelophysis. Dr. James Kirkland opined that it was similar to Nedcolbertia, a small coelurosaur found in Utah. The problem here is that no one knows much more about Nedcolbertia either and its relationships to other dinosaurs are unclear. Quinn and Colbert thought it may have been an ornithomimid, but closer inspection by Rebecca Hunt-Foster and comparison with known ornithomimids indicates this is unlikely. Right now, all that can really be said is that it is likely a small coelurosaur, but not a tyrannosaurid, ornithomimid, or advanced form more closely related to birds, which leaves a small group of poorly known coelurosaurs no one really knows what to do with.
Even if we don’t know for sure what it is, we do have some clues and can narrow down, at least a little, what it might be. What we know for sure is that it is some kind of coelurosaur. That, unfortunately, doesn’t help us a lot because coelurosaurs cover everything from little compsognathids to giant tyrannosaurs to modern birds, known principally for having bigger brains than earlier theropods, slender feet with three toes, and many of them had feathers. It does tell us it is not closely related to dinosaurs like allosaurs and spinosaurs, nor to early theropods like ceratosaurs and Coelophysis. Dr. James Kirkland opined that it was similar to Nedcolbertia, a small coelurosaur found in Utah. The problem here is that no one knows much more about Nedcolbertia either and its relationships to other dinosaurs are unclear. Quinn and Colbert thought it may have been an ornithomimid, but closer inspection by Rebecca Hunt-Foster and comparison with known ornithomimids indicates this is unlikely. Right now, all that can really be said is that it is likely a small coelurosaur, but not a tyrannosaurid, ornithomimid, or advanced form more closely related to birds, which leaves a small group of poorly known coelurosaurs no one really knows what to do with.
Using these animals as a comparison, what can we say about what kind of animal “Arkansaurus” was? It was likely a fast runner with probably an omnivorous diet, eating smaller animals and supplementing its diet with plants. It would likely have stood somewhere between 2-4 meters (6.5-13 feet) tall. It would have looked something like an ostrich with long arms ending in hands with three functional fingers, with one of them being at least semi-opposable, and a jaw filled with small teeth. If it had feathers (which seems increasingly likely), the feathers would have looked more like fur than the large feathery plumage seen on ostriches today. It would also have had large eyes like ostriches, with excellent color vision, based on the fact that its nearest living relatives, crocodilians and birds, all see a broad spectrum of colors (even better than humans).
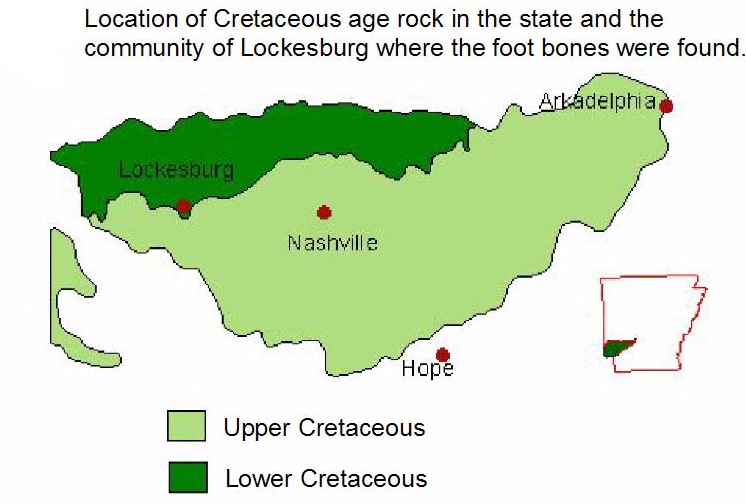 The rocks the bones were found in were part of what is called the Trinity Group. These rock layers (or strata) consist of layers of sand, clay, gravel, limestone, and gypsum laid down in the Early Cretaceous Period, roughly around 100-120 million years ago (what is known as the Albian and Aptian Ages). The rocks indicate that during the time the rocks were formed, the environment was a shallow marine coastal area not unlike south Texas near the Rio Grande or in the Persian Gulf. Our dinosaur would certainly not have been alone. There were other dinosaurs in the vicinity, we just know very little about them. Sauropods left thousands of tracks in the coastal sediment forming a massive trackway found in a Howard Country gypsum mine in 1983. Another trackway found in 2011 has tracks from sauropods such as Pleurocoelus and Paluxysaurus (which may or may not refer to the same species and may or may not also be called Sauroposeidon) as well as tracks from what was probably the giant theropod Acrocanthosaurus.
The rocks the bones were found in were part of what is called the Trinity Group. These rock layers (or strata) consist of layers of sand, clay, gravel, limestone, and gypsum laid down in the Early Cretaceous Period, roughly around 100-120 million years ago (what is known as the Albian and Aptian Ages). The rocks indicate that during the time the rocks were formed, the environment was a shallow marine coastal area not unlike south Texas near the Rio Grande or in the Persian Gulf. Our dinosaur would certainly not have been alone. There were other dinosaurs in the vicinity, we just know very little about them. Sauropods left thousands of tracks in the coastal sediment forming a massive trackway found in a Howard Country gypsum mine in 1983. Another trackway found in 2011 has tracks from sauropods such as Pleurocoelus and Paluxysaurus (which may or may not refer to the same species and may or may not also be called Sauroposeidon) as well as tracks from what was probably the giant theropod Acrocanthosaurus.

Howard County trackway.
Most of the information and images in this post not directly linked to came from the following sources. Many thanks to Rebecca Hunt-Foster for clean pictures from her paper, which she also graciously supplied.
Hunt, ReBecca K., Daniel Chure, and Leo Carson Davis. “An Early Cretaceous Theropod Foot from Southwestern Arkansas.”Proceedings Journal of the Arkansas Undergraduate Research Conference 10 (2003): 87–103.
Braden, Angela K. The Arkansas Dinosaur “Arkansaurus fridayi”. Little Rock: Arkansas Geological Commission, 1998.
The top image is a Leo Cate T. rex. Photo by Debra Jane Seltzer, RoadsideArchitecture.com.
UPDATE: Arkansaurus has recently been named the Arkansas official state dinosaur, reviving interest in the fossil. It is currently being re-examined by Dr. Rebecca Hunt-Foster, with the hopes that new fossils and information that has come to light since her last publication will provide a more refined determination of its relationships.

