Home » Mystery Monday (Page 4)
Category Archives: Mystery Monday
Stepping into a Belated Fossil Friday
Were you able to figure out what last Monday’s fossil was? It is on display at Mid-America Museum in Hot Springs, AR. Ordinarily this would have been posted last Friday, but real life intervened. Apologies for that. Part of what happened was that when I posted the original picture last Monday, I thought I understood the background behind the fossil. It turns out that new research was published in 2013 that changed a lot of the more detailed interpretations. It didn’t change anything of importance to anyone not obsessed with details, but it sent me on a three day search for answers.

What we are looking at here is a foot print of a sauropod. Sauropods were herbivorous, long-necked dinosaurs and were the biggest animals to ever walk the earth, some of them possibly massing 50-80 tons and stretching well over 30 m (100 feet). We can’t say exactly which one made this particular footprint, but we can take a pretty good guess. If you guessed Sauroposeidon, or Astrodon, or Pleurocoelus, or Paluxysaurus, or Astrophocaudia, or Cedarosaurus, you are at least partially correct. These are all titanosaurs, a subgroup of sauropods. But which one we call it is more problematic. It is usually almost impossible to tell exactly which species made a particular track and in this case, it gets even harder because there isn’t a lot of agreement over which names are even valid.

 Before we get into that morass, what is a titanosaur anyway? Titanosaurs have been in the news recently with the discovery of Dreadnoughtus. Most people are familiar with Diplodocus and Brachiosaurus, the two iconic sauropods. These two dinosaurs are the best known representatives of the two main groups of sauropods, with many species in each group. Diplodocus had shorter front legs than back legs and was relatively thin with a long, whip-like tail. It’s head was small and elongate, with simple, peg-like teeth in the front of the jaws. Brachiosaurus had longer legs in front than in back and was stockier, with a shorter, stubbier tail. It’s head was larger, with spoon-shaped teeth. Titanosaurs had front legs that were roughly the same length as the back legs, with a relatively whip-like tail like Diplodocus, although not thought to be as long. The heads looked like Brachiosaurus, but more elongate. Some had teeth like Diplodocus, some like Brachiosaurus. Basically, if you try to envision an intermediate form between Brachiosaurus and Diplodocus, you would wind up with something that looked like a titanosaur, which is rather interesting because all the studies trying to figure out their relationships place titanosaurs as much more closely related to brachiosaurs than to diplodocids. In fact, titanosaurs likely evolved from early brachiosaurids, which means that all the characteristics that make them look sort of like diplodocids are examples of convergent evolution, if the hypotheses about their relationships are correct.
Before we get into that morass, what is a titanosaur anyway? Titanosaurs have been in the news recently with the discovery of Dreadnoughtus. Most people are familiar with Diplodocus and Brachiosaurus, the two iconic sauropods. These two dinosaurs are the best known representatives of the two main groups of sauropods, with many species in each group. Diplodocus had shorter front legs than back legs and was relatively thin with a long, whip-like tail. It’s head was small and elongate, with simple, peg-like teeth in the front of the jaws. Brachiosaurus had longer legs in front than in back and was stockier, with a shorter, stubbier tail. It’s head was larger, with spoon-shaped teeth. Titanosaurs had front legs that were roughly the same length as the back legs, with a relatively whip-like tail like Diplodocus, although not thought to be as long. The heads looked like Brachiosaurus, but more elongate. Some had teeth like Diplodocus, some like Brachiosaurus. Basically, if you try to envision an intermediate form between Brachiosaurus and Diplodocus, you would wind up with something that looked like a titanosaur, which is rather interesting because all the studies trying to figure out their relationships place titanosaurs as much more closely related to brachiosaurs than to diplodocids. In fact, titanosaurs likely evolved from early brachiosaurids, which means that all the characteristics that make them look sort of like diplodocids are examples of convergent evolution, if the hypotheses about their relationships are correct.

The titanosaur Argentinosaurus. Wikipedia
What’s in a name?
Now that we know basically what we are looking at, what do we call the one which may have made this track? That is an excellent question. Two different trackways have been found in Arkansas, both in a commercial quarry in Howard County. They were fantastic finds, with thousands of tracks (5-10,000 tracks in the first trackway alone), placing them among the biggest dinosaur trackways ever found. Unfortunately, other than a few tracks that were spared, they no longer exist as they were destroyed by the quarry operations. That is a sad loss for paleontology, but in defense of the quarry owners, the tracks were found on private land and the owners had no legal requirement to tell anyone about them at all, they are running a business after all. They allowed scientists to study the trackways and in the case of the second trackway, they approached scientists at the University of Arkansas at Fayetteville about the tracks on their own initiative, giving them the opportunity to study the tracks before they were destroyed. As a result, careful maps were drawn, some tracks were removed and others were saved as casts. So the trackways themselves may be gone, but the knowledge of them is still with us and in the public domain.
The tracks were initially described as being from either Astrodon or Pleurocoelus, based on the fact that fossils from these dinosaurs have been identified in Oklahoma in rock units called the Antlers Formation, which is correlated with the Trinity Formation in southwest Arkansas. However, some researchers have concluded that the material upon which these names are based can not be reliably distinguished from any other titanosaur, so the names are what is called nomen dubium, literally dubious names. Pleuocoelus became what is commonly referred to as a junk taxon, which are used as a waste basket for material not identifiable as something else. In this case, when people found bits of a titanosaur in the southern United States they couldn’t identify, they said, it’s um…uh…Pleurocoelus? Pleurocoelus! Yeah! That’s the ticket! In 2013, Michael D’Emic published his research in which he found that part of the material identified as Pleurocoelus are really from two different sauropods called Cedarosaurus and Astrocaudia, and other parts are from a Texan sauropod called Paluxysaurus, leaving other bits unidentifiable as anything other than indeterminant titanosaur. Additionally, he found that Paluxysaurus was simply a juvenile form of Sauroposeidon, a giant sauropod known from four huge cervical (neck) vertebrae found in Oklahoma. So in conclusion, what can we say about the tracks? They were made by a titanosaurid sauropod.
Life’s a Beach
The first trackway was found by Jeff Pittman in 1983 while he was working in the quarry for his master’s degree at Southern Methodist University (SMU). The second set was found in 2011 by quarry workers, who brought it to the attention of Stephen Boss, a geologist at the University of Arkansas at Fayetteville. The tracks in the first trackway were 12-24″ across and were interpreted as being from from adult sauropods. The other trackway was more diverse, with tridactyl (three-toed) footprints attributed to the giant carnivorous dinosaur Acrocanthosaurus, as well as tetradactyl (four-toed) tracks which may have been made by a crocodilian of some sort. The pictures below are of the first trackway, taken by David Gillette, and can be found at his site discussing Seismosaurus.


The rocks in which both of the trackways were found is in what is called the DeQueen Limestone, a subunit of the Trinity Formation. These rocks were laid down in the Early Cretaceous about 115 to 120 million years ago. At the time, the shore of the Gulf Coast went through Arkansas, so much of southwest Arkansas was underwater. The DeQueen Limestome has thin layers of sandy limestone, many of which are quite fossiliferous, with oyster shells in abundance. There are also layers of limy clay and gypsum, indicating the air was fairly hot and dry. Stephen Boss likens the environment at the time to be similar to the Persian Gulf of today. So what we have is the coast of a very warm shallow sea. The dinosaurs appear to have been using the area as part of a migratory pathway. So while no bones of these dinosaurs have been found in Arkansas yet, we know they were here, so keep an eye out when you are fossil-hunting in southwest Arkansas. Who knows, you might find something bigger than you imagine.

Depiction of the environment during the formation of the trackway. Mark Hallett. http://www.columbia.edu/dlc/cup/gillette/gillette19.html
Mystery Monday
This week’s Mystery Monday fossil is a bit different. See if you can guess what it is.
This one is on display in the state. Bonus if you can guess what type of fossil it is, what animal made this fossil, and where it is displayed. Come back Friday for the answers.
Mystery Monday Returns
Welcome back! the new school year has started for most, if not all, people by now. Everyone is busily trying to figure out new schedules, new curricula, new people, sometimes even new schools. Changes are everywhere this time of year. Paleoaerie is no exception. We didn’t get quite as much done over the summer as we would have liked (does anyone?), but it was an interesting summer, filled with good and bad. To start with the bad, the UALR web design course that was initially going to work on revamping the website is no more due to unexpected shakeups at the school. Nevertheless, a different course will take a look at the site and see what they can do, although they sadly won’t have as much time to deal with it.
 But there was a lot of good that happened. Big news for Paleoaerie is that we are now partnered with the Arkansas STEM Coalition, a nonprofit advocacy group for STEM education within Arkansas. This is really important for us because this means Paleoaerie now operates as an official nonprofit organization. What does this mean for us and you? It means that any donation to the site is tax-deductible. It also means that many grants that we could not apply for before are now within possible reach. Fundraising should be a bit easier from now on, which means we may be able to do much more in the upcoming future. One of the things we will be doing in the near future is a Kickstarter campaign to buy a 3D laser scanner so that we can start adding 3D images of Arkansas fossils onto the website, which will be available for anyone to use. One might ask why not use some of the cheap or even free photographic methods that are available. In a word: resolution. I’ve tried other methods. When one is attempting to make a 3D image of an intricate object only a few centimeters across, they don’t work well. If you want details to show up, you need a better system. Stay tuned for that.
But there was a lot of good that happened. Big news for Paleoaerie is that we are now partnered with the Arkansas STEM Coalition, a nonprofit advocacy group for STEM education within Arkansas. This is really important for us because this means Paleoaerie now operates as an official nonprofit organization. What does this mean for us and you? It means that any donation to the site is tax-deductible. It also means that many grants that we could not apply for before are now within possible reach. Fundraising should be a bit easier from now on, which means we may be able to do much more in the upcoming future. One of the things we will be doing in the near future is a Kickstarter campaign to buy a 3D laser scanner so that we can start adding 3D images of Arkansas fossils onto the website, which will be available for anyone to use. One might ask why not use some of the cheap or even free photographic methods that are available. In a word: resolution. I’ve tried other methods. When one is attempting to make a 3D image of an intricate object only a few centimeters across, they don’t work well. If you want details to show up, you need a better system. Stay tuned for that.
![]() Paleoaerie is also partnering with the Museum of Discovery and the University of Arkansas at Little Rock for a National Fossil Day event on October 11. Make sure to mark your calendars and come out to the museum to see the spectacle and diversity that can be found in Arkansas. There is much more than you think. We are also working with the museum on a new dinosaur traveling exhibit. It is very cool, so watch for it later this fall.
Paleoaerie is also partnering with the Museum of Discovery and the University of Arkansas at Little Rock for a National Fossil Day event on October 11. Make sure to mark your calendars and come out to the museum to see the spectacle and diversity that can be found in Arkansas. There is much more than you think. We are also working with the museum on a new dinosaur traveling exhibit. It is very cool, so watch for it later this fall.
The last big news that happened recently is today’s Mystery Monday fossil. Someone brought me a fossil to examine a couple of weeks ago. The first amazing part of it is that is was actually a fossil. the vast majority of what people show me are just interestingly shaped rocks. This was a bona fide fossil. Not only was it a fossil, but a really cool one. The image below is a vertebra from a little seen animal in Arkansas and not at all for a very long time. The fossil is roughly 100 million years old, putting it in the Cretaceous Period. At that time, Arkansas was on the shoreline of the late Cretaceous Interior Seaway. Take a look at the image below and see if you can figure out what it came from. I’ll let you know what it is Friday. Thanks to Matt Smith for bringing this wonderful fossil to my attention. Come out to the National Fossil Day event and see it for yourself.

Taphonomy Tuesday
 Taphonomy Tuesday? What the heck is taphonomy, you ask? Taphonomy is the study of burial processes and all the changes that take place betwixt death and being collected as a fossil. One might also include the effect of sexiness in what fossils get studied and forgotten about, way more people are interested in tyrannosaurs than fossil mosquitos, for instance. Change comes to all things and paleoaerie is no exception. You may have noticed that the answer to Monday’s mystery fossil did not get posted on the blog last week, although it was posted on the Facebook page. That will be rectified today. But first, a couple of pieces of news.
Taphonomy Tuesday? What the heck is taphonomy, you ask? Taphonomy is the study of burial processes and all the changes that take place betwixt death and being collected as a fossil. One might also include the effect of sexiness in what fossils get studied and forgotten about, way more people are interested in tyrannosaurs than fossil mosquitos, for instance. Change comes to all things and paleoaerie is no exception. You may have noticed that the answer to Monday’s mystery fossil did not get posted on the blog last week, although it was posted on the Facebook page. That will be rectified today. But first, a couple of pieces of news.
Mystery Monday and Fossil Friday will be suspended for the summer so I can work on other aspects of the website. I would like to make some different posts and add more to the site. however, for those of you who are primarily interested in the fossils, they will return along with school, teachers, and homework, although hopefully a bit more fun than either doing or, even worse, grading homework (trust me, having been both a student and teacher, grading is almost always more painful than doing the assignments). In the meantime, you can look forward to more varied posts, the addition of a couple of things, such as a techno page for recommended apps and multimedia and an Amazon store in which you can peruse recommended items (and in a small way support the work of paleoaerie, which while free to you, is not to me). So I hope you will stick with me through these Darwinian changes and avoid the pitchforks and torches (ed. note: in keeping with the medieval theme of the picture, I thought about saying burning faggots, but so few people these days know that in olden times, faggot simply meant a bundle of sticks, language too evolves).
 The other big news is that I have started a collaboration with the Museum of Discovery in Little Rock to host a celebration of National Fossil Day in October. I am really looking forward to it and, between me and the museum educators at the MoD, we have a lot of ideas for things to do. I hope everyone can come out and enjoy the festivities and learn about fossils in Arkansas and beyond firsthand.
The other big news is that I have started a collaboration with the Museum of Discovery in Little Rock to host a celebration of National Fossil Day in October. I am really looking forward to it and, between me and the museum educators at the MoD, we have a lot of ideas for things to do. I hope everyone can come out and enjoy the festivities and learn about fossils in Arkansas and beyond firsthand.

Platecarpus skull fossil at Plano West Senior High Photo Credit:Wesley Kirpach. Wikimedia Commons.
Now, for that fossil…Did any of you recognize this as the skull of a mosaaur, specifically that of Platecarpus? To picture a mosasaur, imagine a komodo dragon, replace the feet with flippers, and compress the tail so it is taller than it is wide (aka laterally compressed) so it looks more fish-like than lizard-like, and you have a pretty good view of a mosasaur. There is a reason for that. Komodo dragons are part of a group of lizard called monitor lizards, which are thought to be close relatives of mosasaurs and so are likely an excellent model for what the ancestral animal of mosasaurs looked like before they became aquatic.

Platecarpus. Wikimedia.
Platecarpus was a carnivorous marine reptile that swam in the Cretaceous seas. While dinosaurs like Tyrannosaurus rex and Triceratops roamed the land, Platecarpus and its relatives patrolled the oceans. This is one of the most common mosasaurs, so much of what we know about them comes from fossil of this genus. It was smaller than many of the other mosasaurs. Some, like Kronosaurus, could reach up to 17 meters, but Platecarpus only averaged around 4-7 meters. Its name means “flat wrist,” alluding to the flippers, although it hardly distinguishes them from other mosasaurs in this regard. What did make them stand out from the other mosasaurs was a relatively shorter snout with eyes that faced more forward, so it probably had better stereoscopic vision, that is, it had better depth perception than most others of its kind. This may be why it had a shorter snout, to prevent the snout from blocking its field of view. Like other mosasaurs, Platecarpus had two rows of teeth on its palatine bones, forming the roof of its mouth. This arrangement actually isn’t all that unusual in lizards and snakes, it is really common in fish. The teeth (all of them, not just the palatal teeth) were pointed and conical, although not as sharp as some of its kin, indicating it went after small, soft prey, like small fish and soft invertebrates like squid or perhaps even jellyfish. They could have gone after larger prey like crocodilians do, but unlike crocodilians which have a strong skull capable of withstanding the forces of ripping a prey item apart, Platecarpus had a much weaker skull which would likely not have stood up to the stresses of the crocodilian death roll (this is when they grab a limb and spin until the limb is ripped off, the moral of the story is of course, never dance with a gator). The overall shape of Platecarpus is stockier than most mosasaurs. This would have had the effect of decreasing surface area relative to body weight, which could have increased its metabolism by holding in heat better.

KUVP1001. Mike Everhart. http://www.oceansofkansas.com
One of the things that makes Platecarpus as a genus so interesting is the fossils that have been found with soft tissue preservation. In one, along with parts of the skin, parts of the trachea (windpipe) were preserved. Originally, they were interpreted as part of a dorsal fin, thus all the early pictures of mosasaurs with fins along their backs. However, it was quickly discovered what the traces really were. To his credit, Williston, the scientist who had reported in 1899 the traces as a dorsal fins was the one who published another short paper three years later saying he had made a mistake and the fossil really showed the cartilaginous rings found in trachea. The tracheal rings have also been interpreted as showing the branching point between the two lungs, which is important because it answered a question about their origins. Every researcher agrees that mosasaurs are lepidosaurs, the group including lizards and snakes, but what wasn’t known was whether or not mosasaurs were derived from aquatic lizards or from snakes. The discovery of a trachea showing two lungs confirms the origin within lizards (snakes only have one lung). It may still be that snakes evolved from mosasaurs, but that is not very likely.
The other piece of soft tissue that has been found is the outline of the tail showing a lobe on the tail forming a very shark-like tail. Until this time, many people had thought the mosasaur swam with an eel-like motion, but the tail and the deep caudal fins indicate a much faster shark-like swimming motion. This in turn has caused people to reevaluate the view they were slow ambush predators, supporting a more active predatory forager. Whether or not other mosasaurs had this fin is currently unclear, so there may have been specializations within this group not seen in the larger mosasaur family. Another such example is the discovery of what has been interpreted as thicker eardrums, which may have allowed them to dive to deeper depths.

Tail fluke preserved around skeleton. Credit: (top) Photograph (by Johan Lindgren), (centre) sketch indicating the bones and skin structures preserved (by Johan Lindgren), and (bottom) life reconstruction (by Stefan Sølberg) of Prognathodon sp.
The southwestern corner of Arkansas was at the edge of the Late Cretaceous Western Interior Seaway, so we have several fossils of them in places like Clark, Hempstead and Howard counties, although you can find them all over the world in the right type of rocks. You can find them in the Brownstone and Marlbrook Marl Formations. These formations are indicative of warm, shallow seas, much like the Bahamas today, which considering the locations, shouldn’t come as too much of a surprise to people. If you were taking a nice vacation on the warm Cretaceous beaches 80-85 million years ago, you might have tried fleeing into the water to avoid the dinosaurs on the beach, but you would have been no safer in the water.
I would like to thank Rachel Moore, who supplied a lot of the research involved in putting this post together.
Fossil Friday
It’s Friday, time for the answer to Monday’s mystery fossil. Were you able to identify it?
 These fossils are from a fish called Enchodus, the “saber-fanged herring.” Teeth of Enchodus are commonly found in the Cretaceous-aged rocks in southwestern Arkansas, especially near Malvern and Arkadelphia in the Arkadelphia and Marlbrook Marl Formations, up into the Paleocene rocks of the Midway Group a bit farther north. In other places you can even find them in rocks of Eocene age, although you will have much better luck in the Cretaceous rocks. At this time, southern Arkansas was shallow to coastal marine. Go to the Bahamas, imagine Enchodus, mosasaurs, plesiosaurs, and plenty of sharks in the water around the islands and you would have a good picture of the landscape back then. They were abundant at the end of the Mesozoic Era and survived the asteroid impact that rang the death knell for many animals, including the non-avian dinosaurs. But they never regained their prominence as a key member of the marine ecosystem, eventually dying out completely in the Eocene sometime around 40 million years ago (the Eocene lasted from 55 to 34 million years ago).
These fossils are from a fish called Enchodus, the “saber-fanged herring.” Teeth of Enchodus are commonly found in the Cretaceous-aged rocks in southwestern Arkansas, especially near Malvern and Arkadelphia in the Arkadelphia and Marlbrook Marl Formations, up into the Paleocene rocks of the Midway Group a bit farther north. In other places you can even find them in rocks of Eocene age, although you will have much better luck in the Cretaceous rocks. At this time, southern Arkansas was shallow to coastal marine. Go to the Bahamas, imagine Enchodus, mosasaurs, plesiosaurs, and plenty of sharks in the water around the islands and you would have a good picture of the landscape back then. They were abundant at the end of the Mesozoic Era and survived the asteroid impact that rang the death knell for many animals, including the non-avian dinosaurs. But they never regained their prominence as a key member of the marine ecosystem, eventually dying out completely in the Eocene sometime around 40 million years ago (the Eocene lasted from 55 to 34 million years ago).

The fish reached sizes over 1.5 meters, which makes them on the large side, but not really big, considering there were mosasaurs in the same waters that surpassed 10 meters. Still, with fangs longer than 5 cm, they would not have been fun to tangle with. They were clearly effective predators on smaller fish and possibly soft animals like squid. At the same time, fossils have been found showing they were themselves prey for larger predators, such as sharks, the above-mentioned mosasaurs, plesiosaurs, and even flightless seabirds such as Baptornis. Baptornis was a toothed, predatory bird, but as it only reached 1 meter or so, it would have only been able to hunt young Enchodus. So like many of us today, Baptornis was always up for a good fish fry.

Hesperornis, a larger version of Baptornis, and a few Enchodus feeding on some herring-type fish. Picture by Craig Dylke.
Enchodus is often called the “saber-fanged herring,” although it is unrelated to herrings. So what then was it? Herrings are what is known as forage fish, meaning they are mostly prey items of larger fish and other animals. Most of the fish called herrings are in the family Clupeidae in the groups Clupeiformes, which includes such well-known fish as sardines and anchovies. Enchodus, on the other hand, has been placed into the group Salmoniformes, which includes trout, char, and of course, salmon. When one typically thinks of trout and salmon, one doesn’t think of bait fish, they think of the fish that eat the bait fish. Thus, Enchodus would better better described as a fanged salmon (they were a bit large to call them fanged trout).
The answer to last week’s Mystery Monday
The answer to last week’s Mystery Monday fossil was supposed to be posted last Friday, which didn’t get done. I am going to have to make some changes in the schedule or change the way I do things because I simply don’t have the time to post a new fossil and give a full discussion of it every week and do anything else. So if you have any suggestions on how you think changes would be best done, let me know. I could cut back to every two weeks; still give a fossil every week, but not go into much discussion of what it is each week; or some other possibility. Let me know your preferences.
At any rate, for the last Mystery Monday fossil, I posted this little fossil.

What we have here is a little goniatite ammonoid that has been pyritized, meaning that the shell has been replaced with pyrite. This type of fossilization is pretty common. As the bacteria eat the organism, some of them will release sulfur, which then combines with the hydrogen in the water to form hydrogen sulfide. When it precipitates out of the water, it usually does so as pyrite. In some cases, like in this one, the pyrite crystals can replace the organism so well that it makes a detailed copy of the original. For obvious reasons, this type of fossilization is called replacement, in which the original oranism is replaced with a mineral, be it calcium carbonate, iron, opal, quartz, or in this case, pyrite.
So what are goniatite ammonoids? Ammonoids are part of the group (often called phylum, but for various reasons the specific rank of the group is often no longer used) Mollusca; which includes snails, (gastropods), clams (Bivalvia, meaning two shells, or less commonly, Pelecypoda, meaning hatchet foot), and the Cephalopods.

Cephalopod diversity. Tolweb.org

Nautilus. Arkive.com
Cephalopods include the squids and octopuses, as well as the Nautilus, which is what concerns us here. If you aren’t familiar with a nautilus, think of a squid inside a spiral shell. Squids used to have shells, either long, straight ones or curved and coiled ones. The only one left of these shelled squid is the nautilus. However, you can still see the remnant of the shell in an internal structure called a squid pen, or in the case of cuttlefish, the cuttlebone. In either case, they are the last vestiges of the external shells we see in the nautilus and the ammonoids.

Squid anatomy. Biologycorner.com
During the Paleozoic and Mesozoic Eras, ammonoids were much more common and much more diverse. During the Jurassic and Cretaceous Periods during the Mesozoic, the type of ammonoids that were most common were the more familiar fossils known as ammonites. Goniatites were much earlier and lived during the Paleozoic.

Nautilus shell. Wikimedia.
How does one tell the difference between the different types of ammonoids? Look at the internal partitions. These partitions, called septae, separate the chambers within the shell. As the animal grows, it adds material to the edge of the shell, making it larger and larger at that end. Once the shell gets big enough, the animal will create a new partition in the back, separating the current body chamber from the earlier, smaller one. A small hole is left in the septae so the siphuncle, a thin tube, can pass through, connecting all the previous chambers. The siphuncle could then be used to pump water or gas in and out of the chambers so they could be used as ballast, allowing fine control of their buoyancy.
 The septae in the nautiloids (the group of ammonoids containing the modern nautilus) are all very smooth, forming a nice curve. The nautiloid septae curve inwards, whereas the ammonoids curve outwards to some extent. Ammonoid septae are also much more complex. Goniatites, the earlier forms, had simple wavy septae. Ceratitic ammonoids created septae in which the waves were more jagged, with what often looks like little saw-toothed crenulations. The later ammonites (many people call all of them ammonites, but this term more properly only refers to the more derived subset) had very complex septae, showing several smaller wave patterns overlaid upon the larger wave.
The septae in the nautiloids (the group of ammonoids containing the modern nautilus) are all very smooth, forming a nice curve. The nautiloid septae curve inwards, whereas the ammonoids curve outwards to some extent. Ammonoid septae are also much more complex. Goniatites, the earlier forms, had simple wavy septae. Ceratitic ammonoids created septae in which the waves were more jagged, with what often looks like little saw-toothed crenulations. The later ammonites (many people call all of them ammonites, but this term more properly only refers to the more derived subset) had very complex septae, showing several smaller wave patterns overlaid upon the larger wave.

Ammonoids were very common in the Paleozoic and Mesozoic and were found throughout the oceans of the time. They typically lived in shallow marine environments all over the world. They also evolved rapidly, so new species tended to appear and disappear on a fairly regular basis. This abundance, diversity, and rapid turnover make them prime index fossils. Index fossils are those fossils which are useful for dating the rock layer and correlating the layer from one spot to another. Using index fossils allows us to piece together a complete sequence of events even if there is no one place that has the entire sequence of rocks preserved. As a result, any fossils that can be used as index fossils become very important to people trying to figure out the history of life on earth, such as this little ammonoid. If you want to find one for yourself, look in almost any of the limestone or chert formations in the Ozark mountains. There are plenty.
Taking a Bite Out of Fossil Friday
My apologies for getting this out so late. But now that we’re here, could you tell what the mystery fossil for this week was? It was a very unusual creature. 

Peafowl, male and female. Wikimedia.
The creature in the picture is a representative of an early shark called Falcatus. It was a very strange shark, in which the males had a forward-pointing spike on its head. This is one of the earliest examples of sexual dimorphism in the fossil record. Sexual dimorphism is when the male and females look different. Humans are sexually dimorphic in a variety of ways, but the classic example is the peacock with its extravagant tail. They are so dimorphic that many people do not even realize the term peacock only applies to the male of the peafowl species. The females are called peahens.

Falcatus reconstructed. Wikimedia.

Ozarcus. The fossil itself. AMNH.
Falcatus was chosen this week because of an interesting shark fossil found in 325 million year old limestone near Leslie, AR. The fossil was recently published and got a lot of press. Pictures of the fossil itself show what appears to be little more than a couple of lumpy concretions stuck together. There is little there that resembles anything like a shark. At least, until you look inside. A concretion is an inorganic structure consisting of layers of minerals that have been precipitated around a central core, much like a rocky onion. Oftentimes, a fossil lies at that core and served as the basis upon which the mineral precipitation got started. So one never knows what one will find cracking open a concretion. It may be nothing more than concentric mineral bands, or it may be an exquisite fossil preserved for the ages. In this case, the scientists got lucky. Not only was there an exquisite fossil, but one in which few had ever seen before: the skeletal structure of an early shark’s gill basket.
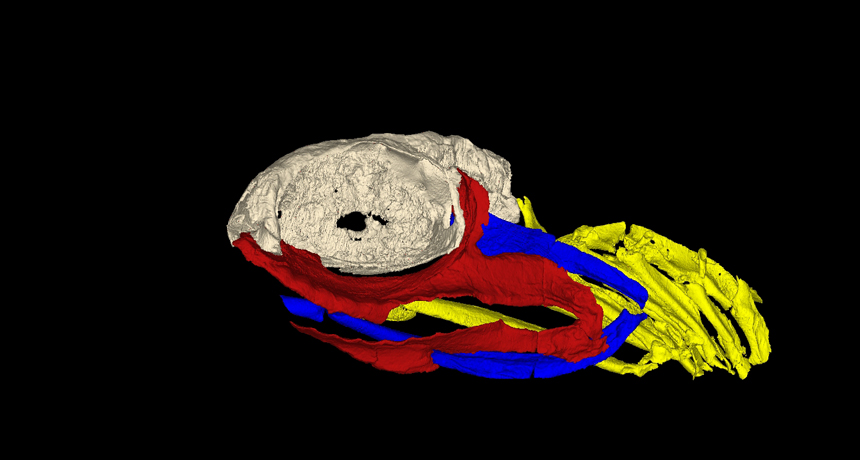
A 3D reconstruction of the skull of Ozarcus mapesae. The braincase is shown in light grey, the jaw is shown in red, the hyoid arch is shown in blue, and the gill arches are shown in yellow. ©AMNH/A. Pradel
The researchers named the fossil Ozarcus mapesae, for the Ozarks in which it was found and for Royal and Gene Mapes, geology professors at Ohio University. The Mapes are also paleontologists and have collected a large number of fossils, one of which happened to be this curious-looking concretion. Small teeth on the exterior pointed to possibly more interesting material inside, so they CT-scanned it, which revealed the remains of the head of the shark. But to get better detail, they had to use a synchotron. This technology is new and expensive enough that it was not possible to look at the fossil this way until recently (a good example of why we preserve fossils in museums, you never know when future technology will allow examination in ways never thought of before).
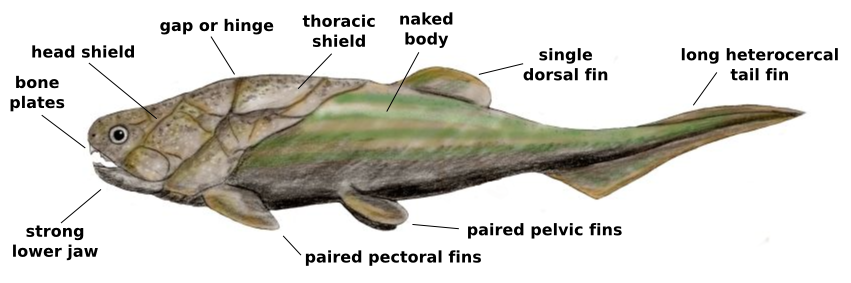
Placoderm anatomy. Wikimedia.
The gill basket goes by many names, the branchial basket, branchial arches, gill arches, etc. but they all refer to the skeletal supports for the gills. To the shark, they are important for holding up the gills and forming the path for water to flow over the gills so the shark can breathe. For us, they are important because the arches evolved into a variety of structures, primarily the jaws and hyoid, a small bone in the throat to which the back of the tongue is attached. Because of the importance of jaws, the evolution of those structures has been a big topic of interest. The traditional view has been that the first jawed fish, the placoderms, evolved into the early precursors of the sharks, which then evolved into the early bony fish, the osteichthyans. The sharks then were expected to have a more primitive structure than bony fish. This story makes sense, given that the order of appearance in the fossil record pretty much matches what we would expect and the skeletal structures look more primitive in sharks than bony fish. Of course, despite the common view that sharks are relics of a bygone age, they have had over 400 million years to evolve after they split off from bony fish. How likely is it that they would have retained such ancestral characteristics for all this time? To answer that question, we can look at the fossil record to tell us what they were like at the earliest stages.
 The big problem with examining the fossil record of sharks is that sharks are chondricthyans. They do not make bone. Other than the teeth, the skeleton is supported by a simpler set of calcium phosphate crystals. Unlike bone, which has a very structured arrangement of crystals and connective tissue, the bones in sharks are made of cartilage and a haphazard set of disorganized crystals, which fall apart shortly after the animal dies. Thus, finding any fossils of sharks that contain more than the teeth is extremely rare. Finding ones in which everything is still in place is almost impossible. Fortunately, over a long enough period of time, even the almost impossible will happen eventually. That is the thing with large numbers and vast amounts of time, our perception of what is unlikely doesn’t really work. For instance, given a 1 in a million chance that a tweet on Twitter will have something requiring the security team to deal with, that still gives them 500 tweets every single day. That “almost impossible” fossil was found with Ozarcus. This fossil provided our first look at what the throat of a primitive shark actually looked like.
The big problem with examining the fossil record of sharks is that sharks are chondricthyans. They do not make bone. Other than the teeth, the skeleton is supported by a simpler set of calcium phosphate crystals. Unlike bone, which has a very structured arrangement of crystals and connective tissue, the bones in sharks are made of cartilage and a haphazard set of disorganized crystals, which fall apart shortly after the animal dies. Thus, finding any fossils of sharks that contain more than the teeth is extremely rare. Finding ones in which everything is still in place is almost impossible. Fortunately, over a long enough period of time, even the almost impossible will happen eventually. That is the thing with large numbers and vast amounts of time, our perception of what is unlikely doesn’t really work. For instance, given a 1 in a million chance that a tweet on Twitter will have something requiring the security team to deal with, that still gives them 500 tweets every single day. That “almost impossible” fossil was found with Ozarcus. This fossil provided our first look at what the throat of a primitive shark actually looked like.
Let’s have a bit of background to cover what we have known of early jaw evolution up to this point. Placoderms, armored fish from the early Paleozoic Era, were the first animals with jaws. The jaws themselves appear to develop from the first gill arch, according to a lot of embryological studies on modern animals. 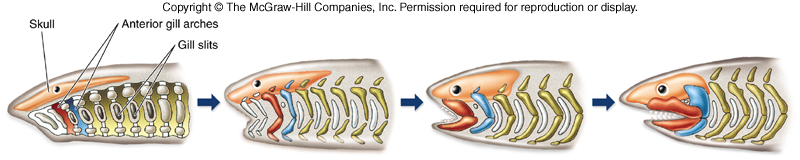
The studies haven’t really answered where the bone came from though. In placoderms, it is pretty clear the armor came from modifications of the dermis, the basal layer of the skin. But they also have internal bone forming their skeleton. Modern sharks have no bone other than teeth and bony fish have jaws made from that dermal bone. In addition to the origins of the bone, there is the matter of how the bones are attached to the skull. The upper jaw is formed by a embryological structure called the palatoquadrate (the top part of the first gill arch), so named because bones called the palatine and quadrate form from it. The bottom jaw forms from what is called Meckel’s cartilage (the bottom part of the first gill arch). In modern fish, the palatoquadrate is braced against the skull only at the front, with the back unattached. The jaw joint itself is attached to a bone called the hyomandibular, which forms from the second gill arch. The hyomandibular acts like a swinging pivot, allowing the jaw to open very wide. When the jaw joint gets pulled forward, the back of the upper jaw can move down, using the point where the front part is braced as the pivot point. Sharks take this to an extreme, not bracing the upper jaw on anything at all, with the jaws attached solely by the hyomandibular bone (the “hyostylic” joint) allowing both the upper and lower jaw to move forward when they open their mouths. So when you see that shark opening its jaws and it looks like they are coming right out at you, they really are. For a great example of a hyostylic jaw joint, check out the goblin shark.
Unfortunately, when we look at the earliest fossil sharks and bony fish, both of them show a jaw in which the upper jaw is braced against the skull in both front and back (the “amphistylic” joint). So it doesn’t tell us much about how the sharks fit into the sequence. One might say that it seems logical to think the bony fish came first, loosening the jaw in the back and then the chondrichthyans took this one step farther. But surely, others might say, the fact that bony osteichthyans have a more advanced bony skeletal structure means they would have to have come later, right? Here again, the fossil record doesn’t help here because the earliest representatives of both groups appear close enough in time that it cannot be strongly stated which came first.
That is where things were, until 2013, when the fossil record started answering these questions. A placoderm called Entelognathus was published in 2013. This fossil was of great interest because it showed the structure of the jaws in great detail. Entelognathus had a jaw that looked very much like an osteichthyan. This fossil was 419 million years old, so it was likely too late to be ancestral to either chondrichthyans or osteichthyans, but old enough to be very close to the ancestral form. What Entelognathus tells us is that the bones forming the jaws in placoderms was already like those seen in modern bony fish, indicating that sharks would have started out with bone, but lost it during their early evolution. Of course, since sharks don’t have bone, this was hard to demonstrate on sharks, so the question was still unsettled.
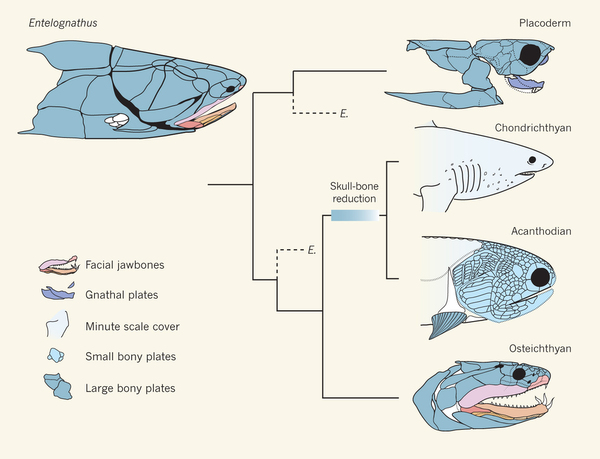
What Entelognathus says about fish relationships. Palaeontology: A jaw-dropping fossil fish. Matt Friedman & Martin D. Brazeau Nature 502, 175–177 (10 October 2013) doi:10.1038/nature12690
Outside of the actual jaws themselves, there are all the other support structures around the jaw, such as the hyoid bone. Many of these structures are formed from the next few gill arches (earlier jawless fish had at least seven gill arches, so using a few to make the jaws and throat still leaves plenty for gills). The skeletal structures in bony fish that support the gill arches form a fairly simple chevron, or wide v shape, whereas the sharks have a slightly more complicated structure. But if finding fossils of shark skeletal structures is rare, finding one with tiny gill arch supports still in position is almost impossible. There is where Ozarcus comes in, because it is here that the almost impossible becomes reality. Once the researchers were able to see inside the fossil with the synchotron, they were able to see preserved gill arch supports in position. That position resembled the simple chevron shape of modern osteichthyans.
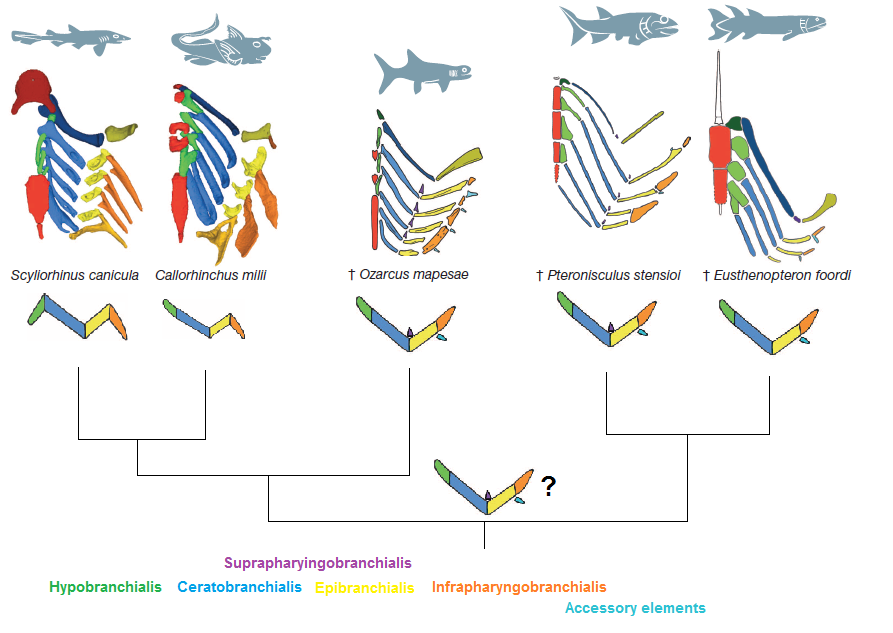
Gill arch supports in Ozarcus are similar to bony fish (right) and not the condrichythyans (left). Pradel A., Maisey J.G., Tafforeau P., Mapes R.H. and Mallatt J. (2014) A Palaeozoic shark with osteichthyan-like branchial arches. Nature DOI: 10.1038/nature13195
Thus, between Entelognathus and Ozarcus, we can confidently assess the development of jaws as having started in placoderms with primitive, osteichthyan jaws. We have evidence of the bony origins of the jaws from Entelognathus and evidence from the gill arches from Ozarcus, so we now have plenty of evidence to strongly support the claim that the chondrichthyans are not in the evolutionary pathway to modern jaws at all. They are an offshoot that actually lost bone to form a more flexible skeleton for some reason. Whether it was mechanical advantage for their lifestyle, such as increasing flexibility, or a reduction in mineral storage, who can say.
This new view of jaw evolution demonstrates that the common view of sharks as being evolutionary relics is wrong. Far from being primitive, they evolved throughout the millenia to the modern sharks we see today, changing the shape and development of their jaws and throat as they evolved from the primitive condition seen in early bony fish into the efficient predators of today. But then, they have been separated from the lineage that led through bony fish up through the early tetrapods all the way to us for over 400 million years. Why would anyone think that in all that time, they stayed still evolutionarily? Even if they look much more similar to their ancestors than we do, they have evolved too. Nothing stays still.
Mystery Monday Fossil
Today’s Mystery Monday fossil is related to some recent paleontology news that I am excited to talk about here because it has put Arkansas fossils on the map in a much bigger way than ever before. See if you can figure out what we have here.
Fossil Friday
On Monday the fossil shown below was put up. How did you do? We got some very good suggestions about it possibly being a horn or a tooth. It isn’t a horn, not exactly. It does also bear a remarkable resemblance to some spinosaur teeth I’ve seen, which was one of the guesses. But it’s not a tooth either. Congratulations to Laurenwritesscience, who really knows her fossils for correctly identifying it as a horn coral.
 Modern day coral are pretty much all in the group Scleractinia, meaning stony rays, known as the stony corals because they make an external skeleton out of aragonite, a form of calcium carbonate, very similar to calcite. They are the major reef builders in modern oceans. But it wasn’t always that way. Before scleractinians, there were tabulate corals and rugose, or wrinkly corals. Rugose corals, like modern corals formed both large colonies and solitary structures. The solitary rugose corals, like this one, are called horn corals, obviously for their commonly stated similarity to horns.
Modern day coral are pretty much all in the group Scleractinia, meaning stony rays, known as the stony corals because they make an external skeleton out of aragonite, a form of calcium carbonate, very similar to calcite. They are the major reef builders in modern oceans. But it wasn’t always that way. Before scleractinians, there were tabulate corals and rugose, or wrinkly corals. Rugose corals, like modern corals formed both large colonies and solitary structures. The solitary rugose corals, like this one, are called horn corals, obviously for their commonly stated similarity to horns.
Rugose corals first appeared in the Ordovician Period roughly 47o million years ago, give or take 20 million years. They became important reef builders in the Devonian Period and remained a highly visible presence until the end of the Permian about 250 million years ago. They, like almost everything else, succumbed during the Great Dying, as the mass extinction of that time is called. As a result, if you want to find them in Arkansas, you should try the marine limestones in northern Arkansas, throughout the Ozarks. The best place to find them are the Mississippian rocks, such as the St. Joe, Boone, and Pitkin Formations.

Life cycle of a rugose coral. Picture from the Kansas Geological Survey
Rugose corals, like all corals, are more than just the stony skeleton most people think of when they see coral. Inside, the animals themselves grow as polyps sitting, in the case of horn corals, a stony cup called a calyx, which they constantly build up as they grow. When alive, the pointy end went down, permanently attached to the surface upon which it grew. Inside the calyx, they grow septae, little walls used to provide structural support for the skeleton and to hang themselves on (while this may cause one to picture a very depressed polyp, a better visual image would be hanging pictures or cabinets on a wall). Rugose corals typically grew six main septae, forming six compartments. Four of the compartments were further subdivided by minor septae. All of these septae radiated from a central axis, or rod supporting the whole structure. The four main compartments is one reason why they are sometimes called Tetracorallia.

Growth of septae in rugose and hexacoral. Paleo.cortland.edu.
This is one of the key characteristics that separate them from modern day coral, which are members of a group called Hexacorallia, noted for having six main septae and six main compartments, all of which are furhter subdivided with minor septae, thus having six-fold symmetry (you can draw six lines through the center of the organism in which one side is a mirror image of the other). Hexacorallia also include the sea anemones, also noted for having six lines of symmetry. The Rugosa, on the other hand, due to the four compartments, often exhibited only bilateral symmetry, meaning there was only one plane you could draw to find a mirror image.

Coral polyp anatomy. Fossilmuseum.net.
The polyp forming the living organism was a tiny creature with a simple sac-like digestive tract, meaning there is only one opening. Food passes into the mouth and waste comes right back out the same way. This is the typical pattern seen in almost all of the Cnidarians, the group in which corals are a part, along with animals like the jellyfish. Like the jellyfish, coral polyps have nematocysts, stinging cells they use to capture prey. Surrounding the mouth, and where one would find the nematocysts, are a ring of tentacles, forming a flower appearance, the reason the group containing all corals is called Anthozoa, the flower animals. A lot of group names and you might be getting lost, so I’ll summarize the names: Horn corals are a part of the Rugosa, or rugose corals, also called Tetracorallia, which are a part of Anthozoa, along with all the other corals and sea anemones, which are a part of Cnidaria, which also includes the jellyfish among others. The modern corals are mostly scleractinians, or stony corals, which are a part of Hexacorallia, which are Anthozoans and thus Cnidarians (which you may also see referred to as coelenterates). Like all members of the Anthozoa, rugose corals almost assuredly reproduced asexually by budding or sexually by releasing both sperm and eggs. In many corals, most of the polyps in an area will do this at or near the same time, called broadcast spawning, creating a massive cloud of eggs and sperm floating around the reef. I have read this is a fascinating sight, if rather disturbing to some. Why do they do this? Because everything else in the reef considers them good eating, so the only way to ensure that some escape long enough to survive and grow is to simply overwhelm them with sheer numbers. This idea is often called predator satiation. It also gets around the fact that coral can’t move, they are the ultimate homebodies. The only hope they have to meet a good mate is to just put the seeds of their offspring into the wind, or in this case, currents, and hope for the best.
Most corals do not depend just on what they can capture or filter out from the water. They also have a mutualistic relationship with photosynthetic dinoflagelates and zooanthellae, or single celled algae. They algae get a safe place to live and nutrients, while the coral gets the products of the photosynthesis, such as oxygen, sugars and amino acids. Normally the coral don’t depend upon the extra energy derived from the photosynthesis, but when times are tough, the added supply can see them through, allowing both the coral and the algae to survive longer. The mutualism certainly provides a much more abundant resource supply for the coral, allowing them to grow more efficiently. We don’t know if rugose coral did this, but it is reasonable to assume they did for now. Of course, this means that corals who use this relationship are restricted to shallow, clear waters conducive to the growth of algae. We do know that they formed close relationships with stromatoporoid sponges. The sponges formed the backbone of the reefs, with the coral growing within the sponge reefs. What sort of association beyond that is not known though.

Life in the Devonian seas. UMich Mus. Natural History.
Mystery Monday

Time for another Mystery Monday fossil! See if you can figure out this wrinkly rock.

