I was reading a post by Brian Switek on his blog Laelops—which by the way, if you aren’t reading it, you should—about interpreting injuries on dinosaur bones. It’s an interesting read, but what caught my eye was a problem that I have seen in more places than I can count. He included this picture in his post. Take at look at the front limbs.

Plateosaurus. Credit: Brian Switek
The “hands” are pointed inwards. To get in this position, the elbows have to be turned outwards, using a rotation in the shoulders. But it also requires the hands to be pronated. To get what I mean by that, hold your hands forward with the palms up. When you do this, the two bones in your forearms—the ulna and the radius—are positioned side by side.
 This position is called supination. Pronation requires the radius to rotate so that it crosses over the ulna. This can be done because of the construction of the elbow. The ulna is essentially the entire elbow joint, making it a hinge type joint. The radius is, for the most part, just along for the ride at the elbow. The head—the part at the elbow—is round, with a shallow indentation, which is surrounded by what is called the annular ligament. That ligament wraps around the radius, attaching it to the ulna, but never actually attaching to the radius itself, allowing the head to spin in the sling. It is that shape of the radius and the annular ligament that allows it to rotate freely, which makes our level of pronation possible.
This position is called supination. Pronation requires the radius to rotate so that it crosses over the ulna. This can be done because of the construction of the elbow. The ulna is essentially the entire elbow joint, making it a hinge type joint. The radius is, for the most part, just along for the ride at the elbow. The head—the part at the elbow—is round, with a shallow indentation, which is surrounded by what is called the annular ligament. That ligament wraps around the radius, attaching it to the ulna, but never actually attaching to the radius itself, allowing the head to spin in the sling. It is that shape of the radius and the annular ligament that allows it to rotate freely, which makes our level of pronation possible.
Importantly, all tetrapods have the same bones. It was set in place from the first fish that developed bones to support their fins and remains that way all through the hundreds of millions of years to us today. However, not all animals have the same shape of the radial head. Some animals appear to not have both bones, but in reality, they do. They have just fused the bones together. But that fusion has consequences, just like altering the shape of the radial head.
Before we move on to dinosaurs and those consequences, it would be reasonable to ask about other animals to see if they show the same pattern. Let’s take a look at proboscideans, the family of elephants. They are large animals that have their palms facing downward.

This is an elephant skeleton on display at the Manchester Museum. They have a radius and ulna, just like humans. Theirs, however, do not swivel. Nevertheless, it is rotated so that the bones are not in parallel, but the two ends are twisted so that the radius is twisted over the ulna. Their forelimbs are in permanent pronation.
So what about dinosaurs? Let’s look at Dreadnaughtus, the giant sauropod.
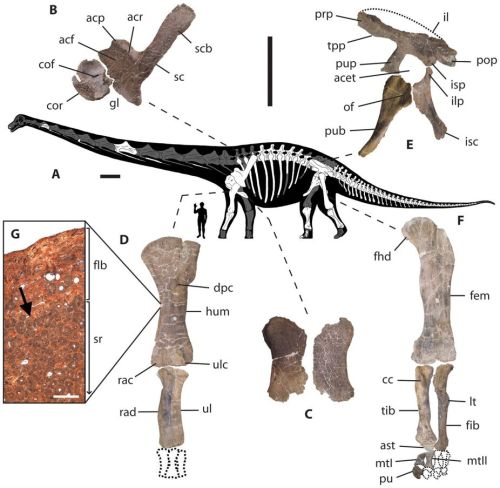
This is Figure 2 from Lacovara et al. 2014. The radius and ulna are massive, as befits a giant quadruped. They are also incapable of rotating to pronate the foot. It has been said by some that some degree of pronation is required for efficient quadrupedal locomotion, but that is not really accurate. It does mean though, that the first digit, what would be our thumb, is going to face forward or at most slightly inward while the remaining digits will be angled outward (anterolaterally). They will not be situated directly forward without shifting the arms at the shoulder. As an example, here is a figure showing a sauropod trackway. Note the direction of the toes.

This is a figure from Falkingham, et al. 2010. As can be seen, the toes are not forward, but pointing outward. The level of pronation is minor and can be achieved with only rotation at the shoulder and a minimal shift or the forelimb. VanBuren and Bonnan (2013) found this was true in all quadrupedal dinosaurs.
This is a best case for pronation in dinosaurs and they can’t do the full pronation needed for bunny hands, much less the bizarre inward facing hands of the Plateosaurus above. So let’s look at theropods, where we see bunny hands all the time. For instance, in Jurassic Park pretty much all the dinosaurs have bunny hands.

The palms are downward with the arms close in, requiring a full pronation to achieve that position. So could they do it? To the bones. This is the radius of Neuquenraptor, as published by Novas and Pol in 2005. It is an unenlagiin, in the family Dromaeosauridae, along with Velociraptor, Deinonychus, and all those other raptor dinosaurs.

These radius is fairly straight, and if you look at the head, it is not the shallowly indented round cup we see in mammals. It is more angular, which is typical of theropods. That relative angularity would prevent the radius from rotating as ours do, in effect locking it into a small range of motion, preventing them from placing their hands palm down.
In point of fact, VanBuren and Bonnan didn’t just look at sauropods. They looked at all types of dinosaurs. They found that no dinosaur had the ability to cross the radius over the ulna, which means that at best, they had very limited ability to pronate their forearms. That means no known dinosaur could have characteristically held their arms close in with their palms facing downward, aka bunny hands.
It is a little more complicated than this. Studies have shown that if the arm is fully extended, the hands can be more pronated, by using the entire length of the arm to rotate, but even that is not going to be fully pronated like we can, or bunnies can. And if you think about the living dinosaurs, the birds, when was the last time you saw a bird put its wings flat on the ground in front of them? They can clap, but they can’t type or play basketball.
Of course, the wrist bones of the maniraptors—those dinosaurs leading up to birds—did have what is called a semilunate carpal bone, allowing them to move their hands to insane degrees side to side, which is what allowed them to develop the ability to fold their wings like they do, and they can flex and extend their hands to a remarkable degree. But they cannot rotate their wrist. Try it yourself. See what movements you can make with your hand without moving your forearm, just your hand. However far you can move your hand that way, these dinosaurs have you beat in spades. But once you move that forearm, you have a serious advantage over them.



Do we know when fore-arm swiveling evolved then? Is it a mammalian trait that elephants lost, or did it pop up in bunnies and primates independently?
It’s a mammalian trait. Elephants didn’t lose it so much as lock it into a pronated position, so it is permanently swiveled, which is pretty common for most mammalian quadrupeds. Dinosaurs never developed it, I expect because they evolved from an at least semi-bipedal and likely an obligate terrestrial biped. Starting from a ground-based biped, which had its hands free may have in a way have set them on a path they never changed for their hands. Mammals, on the other hand, started from small quadrupedal animals that either burrowed or were arboreal. A forelimb that can swivel would be a big benefit for both activities. Being small and rodent-like, they likely had short generation times, which would have given them both the evolutionary time and selective pressure to develop their forelimbs in such a way. The dinosaurs did not seem to have had similar selective pressures to do so. The theropods leading to birds actually had selective pressures to not do so. Instead, they needed to strengthen the arms by locking the forelimb into place, which had the side effect of preventing them from rotating their wrists. They increased the ability to move the hand in other directions instead. The mammals needed forelimbs that could swivel pretty much at the beginning, so mammals developed it essentially as their default position.
When you’re a sprawling terrestrial quaduped, you’re not going to evolve the ability to cross your forearm bones, because the forearm is weaker when it’s crossed than when the bones are parallel. And indeed, such animals as dicynodonts or pareiasaurs didn’t, no matter how vertical their hindlimbs were. The trick is that the origin of therian mammals and their gait with vertical limbs involved a “size squeeze”: tiny animals living in trees, walking on twigs. Forearm mobility is important, weakness is not because they don’t weigh much. Dinosaurs never went through such a phase. Instead of from tiny arboreal quadrupeds, they come from reasonably large terrestrial bipeds.
Sauropods cheat. In the trackway you show, the feet point pretty far outwards, but the hands don’t do that much. And yet, as you correctly point out, the forearm bones don’t cross. That’s because the whole radius, not just its distal end, has moved around the ulna, from a preaxial (cranial, anterior) to a plantar (medial) position – or rather the ulna has grown around the radius, evolving a volar (lateral) lamina and becoming Y-shaped in cross-section in the process. No mammal has done that.
Thanks for the more in depth explanation. I have not spent that much time looking at sauropod forelimbs, so my explanation was rather cursory. I appreciate you filling in the details better on both the sauropods and the additional comments on the early mammals.
https://polldaddy.com/js/rating/rating.js” Instead, they needed to strengthen the arms by locking the forelimb into place, which had the side effect of preventing them from rotating their wrists.”
I’ve wondered about this too. In theropods, the motion of the forelimb is highly constrained. As you say, it might be an adaptation to the rigors of predation – to make the forelimb more resistant to thrashing, struggling prey. But many theropods probably targeted prey much smaller than themselves. Also, many theropods were not carnivores at all (such as oviraptorosaurs).
One possibility is that the large and highly specialized semilunate carpal has nothing to do with predation, but is simply a mechanism to fold up long hands when not in use (and also to protect the attached feathers). I’m skeptical that the swivel-wrist was a side effect of a strong, prey-catching forelimb.
P.S. Great article. 🙂
Thanks, glad you liked it. Although I don’t see any place where I said it had anything to do with predation, only that they needed to strengthen that joint. There are many such potential reasons for that, predation being only one. As David pointed out, dinosaurs never went through a phase prioritizing mobility over strength. In truth, maniraptorans had pressures to increase the arm strength. In regards to the semilunate carpal evolving due to selective pressures having nothing to do with predation, I quite agree. At best, predation is only one causal factor, and possibly, little to none.
Yep, sorry about that – I over-interpreted what you wrote. I agree that there are many potential reasons for strengthening the arms.
I really like David’s point: “Dinosaurs never went through such a phase. Instead of from tiny arboreal quadrupeds, they come from reasonably large terrestrial bipeds.”
And it’s likely that they always remained bipeds. IMHO, the lack of mobility in theropod arms is strong evidence against any theropod being an arboreal quadruped. The forelimbs would have been useless for climbing… even juvenile-hoatzin-style (where the proportionally huge feet do most of the clambering anyway).
David did make a good point. I have to disagree about the arms being useless for arboreality. You say arboreal quadruped, so do you mean to limit it strictly to quadrupedality or arboreality in general? Clearly, there are numerous arboreal birds, so we know they can be arboreal. Ken Dial did a good job showing the utility of even pathetically tiny wings for climbing using his WAIR (wing assisted inclined running). When climbing trunks, most animal do not need to pronate their arms. Rather, they basically hug the tree, with their palms facing inward. Now once they are up in the tree, there are many different ways to get around, which makes determining if an animal is arboreal only from their skeletons a very tricky endeavor. Many different types of animals have figured out arboreality. Mammals are the only ones to use pronation to do it and even they don’t use it to get into the trees. Snakes for instance, don’t even have limbs and they do just fine, as do numerous lizards, birds, and insects. Honestly, as good a point as David made, I don’t think pronation developed due to arboreality. I view it more as a preadaptation that helped them easily become arboreal. I will have to do more checking, but I expect pronation developed in the dicynodonts before arboreality was a factor. I could be completely wrong about that though, so I welcome anyone who knows definitively about that to correct me if I am mistaken.
https://polldaddy.com/js/rating/rating.jsWAIR has the problem that it requires dorsally pointing shoulder joints. Even lateral does not seem to be enough.
And yet it works. I have seen no convincing evidence that maniraptorans were incapable of doing so. Can’t say that I have read everything about it, but the critiques I have read of it were not persuasive to me. They clearly have not been persuasive to Dial and a goodly number of others, so i don’t think my skepticism of the complaints is too out there. Admittedly, I haven’t read any critiques published within the last few years, so maybe there are more viable criticisms out there. I just haven’t seen them.
https://polldaddy.com/js/rating/rating.jsDicynodonts, like crocodiles today, had semi-sprawling forelimbs and could not pronate.
Spent some time looking at dicynodont radii. Seems you are right. Another interesting hypothesis felled by facts. Thanks for bringing that to my attention.
https://polldaddy.com/js/rating/rating.jsWAIR seems to require a fairly sophisticated flight apparatus. David mentioned the orientation of the shoulder joint (glenoid) – that’s one problem. The musculature required for WAIR is also quite demanding. WAIR is more likely to be a derived avian behavior of juvenile birds, not an incipient maniraptoran behavior that could lead to flight. Dececchi et al. (2016) doi: 10.7717/peerj.2159 have a great discussion of this. I don’t think WAIR is a viable pre-flight behavior in the theropod ancestors of birds.
Not sure what you are getting at here. The Dececchi paper does not support your conclusions. It states that WAIR is seen most prevalently in juveniles, but it is seen in all ages in many birds. It is to be expected that it would be seen most commonly in juveniles. Adults can simply fly. I was in the audience when Dial first presented on WAIR. He proved then that birds of all ages did it. The older the bird, the better it performed, which is supported in the Dececchio paper. It does not say WAIR could not be done, only that there was no consistent progression towards it. They in fact demonstrated that derived deinonychosaurs were capable of WAIR. They also showed that none of the tested hypotheses showed any consistent pattern of development in maniraptorans. Far from discounting WAIR, they showed its viability for the most derived deinonychosaurs.
Not sure what you are getting at here. The Dececchi paper does not support your conclusions. It states that WAIR is seen most prevalently in juveniles, but it is seen in all ages in many birds. It is to be expected that it would be seen most commonly in juveniles. Adults can simply fly. I was in the audience when Dial first presented on WAIR. He proved then that birds of all ages did it. The older the bird, the better it performed, which is supported in the Dececchio paper. It does not say WAIR could not be done, only that there was no consistent progression towards it. They in fact demonstrated that derived deinonychosaurs were capable of WAIR. They also showed that none of the tested hypotheses showed any consistent pattern of development in maniraptorans. Far from discounting WAIR, they showed its viability for the most derived deinonychosaurs.
The idea that it requires a sophisticated flight apparatus only applies if one assumes it requires the musculature of modern birds. Dececchio proved that is not true. If we assume it takes modern bird apparatus to fly, much less WAIR, we would be forced to say that bats and pterosaurs could not fly. We can clearly falsify that. David is a smart guy, but I frankly have never understood his disdain for WAIR. I do not agree with him that the shoulder joint needs to be as specialized as he claims. I have seen nothing to support the contention that deinonychosaur shoulders were incapable of WAIR. The only ones I have seen claim that used unrealistic shoulder reconstructions. Gatesy or someone might have published better reconstructions I have missed, but the ones I have seen used unsupportable assumptions, often unsupportable restrictions, and sometimes not sufficient restrictions in other areas because they failed to properly account for soft tissue development. WAIR might be discarded in favor of a better hypothesis in the future, but we are a long way from right now.
https://polldaddy.com/js/rating/rating.js” You say arboreal quadruped, so do you mean to limit it strictly to quadrupedality or arboreality in general? Clearly, there are numerous arboreal birds, so we know they can be arboreal. ”
Yes of course, many birds are arboreal. The question is, for theropods, did arboreality precede flight, or flight precede arboreality? I’d say the latter. Arboreality is not just the ability to climb or clamber up vegetation (including trees). Many animals can do that – goats, crocodiles, even turtles. None of these animals are ‘arboreal’. Arboreality is spending most of the time in trees, and having adaptations for this.
” Now once they are up in the tree, there are many different ways to get around, which makes determining if an animal is arboreal only from their skeletons a very tricky endeavor. ”
I disagree with you here. Determining if an animal is arboreal from the skeleton is not as tricky as it is sometimes made out to be. For example, it is sometimes claimed that tree kangaroos are poorly adapted to living in trees, and that their skeletons do not show clear arboreal adaptations. But this is simply not the case. Tree kangaroos actually possess many arboreal adaptations; but because they are secondarily arboreal macropods, they are not as adept in their arboreality as the related phalangeriform marsupials (primitively arboreal).
The tricky bit is differentiating scansorial and arboreal adaptations, which is harder to do, especially for small mammals.
Deinonychosaurs (including microraptorans) and Archaeopteryx show neither scansorial or arboreal adaptations. Basal pygostylian birds like confuciusornithids and sapeornithids show arboreal (perching) adaptations in the feet, but no scansorial adaptations (including trunk-climbing adaptations). The long, parasagittal hindlimbs and hinge-like (mesotarsal) ankles would have made trunk-climbing extremely difficult. If early birds could launch into the air from the ground (ground-level take-off), which is a real possibility, it would make climbing unnecessary.
Actually, all non-theriimorph synapsids (and maybe some theriimorphs) were incapable of pronation in their forelimbs, because their forelimbs were (semi-)sprawling (even though some had erect hindlimbs, like the afforomentionated dicynodonts, eucynodonts [such as Trucidocynodon and Brasilodon] and most non-monotreme, non-theriimorph mammaliaforms, or at least facultatively erect hindlimbs, like most eutherapsids). Some (or all) theriiforms such as at least some eutriconodonts, multituberculates, Zhangheotherium (representating symmetrodonts), Henkelotherium (representing dryolestoids) and therians were (and are), in contrast, able to pronate, even though all except the last had (semi-)sprawling forelimbs (hindlimbs being either [semi-]sprawling, such as multituberculates, Akidolestes and some eutriconodonts, or erect, like all the others [except fossorial forms such as moles, which have more abducted hindlimbs] ).
The ability to pronate in theriimorphs may be due to a facultatively erect forelimbs in the clade (which became obligatorily erect in Jeholodens and therians, and in most of the others, the shoulder girdle and forelimbs have at least some similarities with therians). Yinotherians, which are the other branch of mammals, in contrast, weren’t (and aren’t) able to pronate.
Also, very interestingly, human arms at rest are in a clapper position like bipedal dinosaurs, although we’re still able to pronate our hands because we very recently evolved from quadrupeds.
There goes my idea of raptors using tools like primates do. Oh, well.
They could probably use tools, just not with the dexterity of primates. But they would likely be more inclined to use their mouth as a sort of third hand as well, since their nack is far more mobile than primates.
There goes my idea of raptors using tools like primates. Oh, well.